人类Y染色体进化历程解读
染色体XY代表男性,XX代表女性,那这个Y染色体是祖祖辈辈留下来的吗?
染色体XY代表男性,XX代表女性,那这个Y染色体是祖祖辈辈留下来的吗?■在大多数情况下,答案是肯定的。
也就是说Y染色体就是从男性祖先那里祖祖辈辈传下来的,而且几乎没有什么改变。
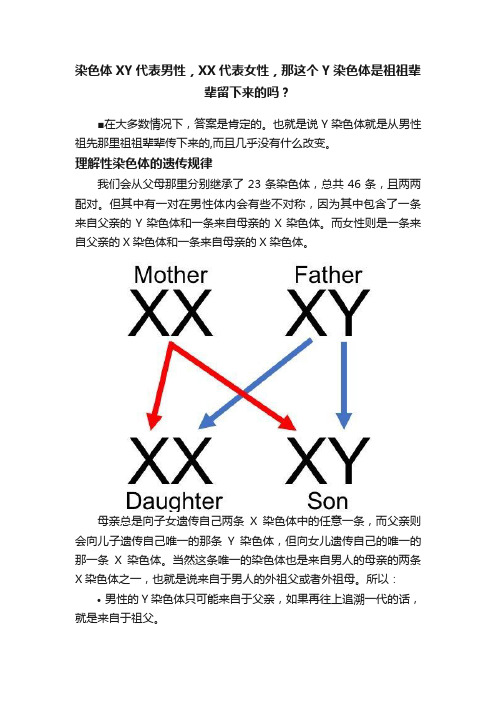
理解性染色体的遗传规律我们会从父母那里分别继承了23条染色体,总共46条,且两两配对。
但其中有一对在男性体内会有些不对称,因为其中包含了一条来自父亲的Y染色体和一条来自母亲的X染色体。
而女性则是一条来自父亲的X染色体和一条来自母亲的X染色体。
母亲总是向子女遗传自己两条X染色体中的任意一条,而父亲则会向儿子遗传自己唯一的那条Y染色体,但向女儿遗传自己的唯一的那一条X染色体。
当然这条唯一的染色体也是来自男人的母亲的两条X染色体之一,也就是说来自于男人的外祖父或者外祖母。
所以:•男性的Y染色体只可能来自于父亲,如果再往上追溯一代的话,就是来自于祖父。
•男性的X染色体只可能来自于母亲,若再往上追溯一代的话则可能来自外祖父或者外祖母。
•女性的两条X染色体,一条来自于父亲,一条来自于母亲,如果再往上追溯一代的话则可能来自于祖母、外祖母或外祖父,但绝对不可能来自于祖父。
上图:X染色体的遗传,男性的X染色体只能来自于母系,女性的染色体则来自父母,但再往上追溯一代不会来自于祖父。
通俗点说,Y染色体永远并且直接来自于爸爸,而X染色体则终归还是来自于某个妈妈(因为男人的X染色体只能妈给)。
在有性生殖的过程中,来自父母的两组染色体将“重组”它们的基因,有点洗牌的意味,这意味着这些染色体彼此交换遗传信息,达到某种随机混合的效果。
从短期来看,这种“洗牌”的过程意味着儿子或女儿的性格和生理特征的组合很难严格复刻父母。
从长远来看,这形成了某种遗传多样性,并有助于消除可能对后代不利的性状。
上图:Y染色体主要基因的功能是涉及性决定以及精子发育。
而X 染色体涉及的问题就多了,诸如眼睛白化症基因、杜氏肌营养不良症、雄激素不敏感症、严重的组合免疫缺陷、血友病、色盲症,与这些疾病相关的基因都在X染色体上。
06人类染色体

Medical Genetics
表6-2 人类染色体分组与形态特征(非显带, Denver体制)
分组 染色体编号 大小
A
1~3
最大
B
4~5
次大
着丝粒位置
中央(1、3号), 亚中央(2号)
Medical Genetics
Medical Genetics
6.1.2.1 染色体四级结构模型
Medical Genetics
6.1.2.2 袢环模型
从DNA到染色体水平的压缩过程
Medical Genetics 6.1.3 染色体形态结构
: 中期染色体的形态特征 姐妹染色单体、着
丝粒(centromere)、短臂(p)、长臂(q)、 端粒、主缢痕、次级缢痕、核仁组织者区、随体
6.2.2.2 染色体高分辨显带
Medical Genetics
6.2.3 分子细胞遗传学技术
6.2.3.1 荧光原位杂交(FISH)
人类BCR、ABL基因双标记荧光原位杂交图
Medical Genetics
6.3 核型识别和细胞遗传学命名原则
6.3.1 非显带核型的识别
核型(karyotype) 一个体细胞中的全部
定义:间期细胞核中能被碱性染料染色的物质,伸 展开的DNA蛋白质纤维。
分类: 常染色质 异染色质
Medical Genetics
常染色质与异染色质的特性比较
特征
常染色质
数量 占染色体的极大部分
分布 位于核中央、核仁内部
染色 正常染色反应,染色浅 复制 正常复制 形态 折叠松散,呈疏松状 组成 含单一和重复DNA序列
分子人类学视角地人类迁徙和种族源流

分子人类学揭示人类迁徙路线和种族源流1、Y-DNA单倍群人类Y染色体DNA单倍群由非重组DNA的Y染色体突变进行定义。
这种由许多人共享的突变称为单核苷酸多态性(SNP)。
人类Y染色体每一代大约积累两次突变。
Y-DNA单倍群的分支结构组成一个Y染色体进化树,有数百甚至数千的突变由这些不同的单倍群共享。
Y染色体的最近的共同祖先(most recent common ancestor MRCA),也被称为Y染色体亚当,是目前活着的男性的最近的男性共同祖先。
Y染色体亚当估计生活在大约236000年前的非洲。
通过研究其他瓶颈,所有欧亚大陆的人都是69000年前的一个男人的后裔。
之后一个主要的遗传瓶颈期发生在大约5000年前,今天大多数欧亚大陆的人可以追溯到5000年前的十二个祖先。
Y-DNA单倍群进化树单倍群A & B现代所有单倍群都是单倍群A的后代,稀疏分布在非洲,主要集中在西南部的科伊桑人和尼罗河谷东北部人群。
单倍群CT (P143)标识单倍群CT的突变标记是M168和M294.包含单倍群D、E、C、F,可能88000年前在亚洲或非洲出现。
单倍群C (M130)•Haplogroup C (M130, M216) 分布在亚洲、大洋洲和北美o Haplogroup C1 (F3393/Z1426)▪Haplogroup C1a (CTS11043)▪Haplogroup C1a1 (M8, M105, M131) 日本低频分布▪Haplogroup C1a2 (V20) 欧洲和尼泊尔低频分布▪Haplogroup C1b (F1370, Z16480)▪Haplogroup C1b1 (AM00694/K281)▪Haplogroup C1b1a (B66/Z16458)▪Haplogroup C1b1a1 (M356) 印度低频分布, 阿拉伯半岛和中国北部▪Haplogroup C1b2 (B477/Z31885)▪Haplogroup C1b2a (M38) 分布在印度尼西亚,新几内亚岛,美拉尼西亚,密克罗尼西亚,和玻利尼西亚▪Haplogroup C1b2b (M347, P309) 澳洲土著o Haplogroup C2 (M217, P44) 分布在欧亚大陆和北美,特别是在蒙古人,哈萨克人,通古斯人,西伯利亚人,和Na-Dené-speaking语民族单倍群D (M174)•Haplogroup D (M174) 分布在日本、中国(西藏)和安达曼岛o D1 (CTS11577)▪D1a (Z27276, Z27283, Z29263)▪Haplogroup D1a1 (M15) 主要分布在西藏、羌族、彝族和苗瑶语人群▪Haplogroup D1a2 (P99) Found mainly in Tibetans, Qiangic peoples, Naxi,and Turkic peoples▪Haplogroup D1b (M55, M57, M64.1, M179, P12, P37.1, P41.1 (M359.1), 12f2.2) 主要在日本o D2 (L1366, L1378, M226.2) 菲律宾Mactan 岛Haplogroup E (M96)[edit]•Haplogroup E (M40, M96) 分布在非洲、中东和欧洲o Haplogroup E1 (P147)▪Haplogroup E1a (M33, M132) formerly E1▪Haplogroup E1b (P177)▪Haplogroup E1b1 (P2, DYS391p); formerly E3▪Haplogroup E1b1a (V38) 非洲尼日尔-刚果语人群; formerly E3a▪Haplogroup E1b1b (M215) 非洲之角,北非、中东和欧洲地中海地区;formerly E3bo Haplogroup E2 (M75)Haplogroup F (M89)单倍群F和后代.单倍群F和后代构成了目前世界人口的90%,几乎都分布在撒哈拉非洲之外。
Investigative Genetics:从Y染色体解析东亚人群历史

Investigative Genetics:从Y染色体解析东亚人群历史作者:王传超李辉来源:科学网2013-6-17 17:14:19分享到:1关键词:东亚人群性染色体东亚是亚洲的一部分,面积广阔、风光旖旎。
东亚有着占世界22%的人口,主要分为四种体质类型:新石器时代东亚人,即蒙古人种;旧石器时代大洋洲人,即澳大利亚人种;旧石器时代东南亚人,即尼格利陀人种;还有欧洲人即高加索人种。
东亚有阿尔泰、南亚、南岛、侗傣、苗瑶、汉藏和印欧等七个语系的200多种语言。
这使得东亚成为世界上研究人类进化、遗传多样性和基因与文化相互作用的最重要区域之一。
在过去数年中,分子人类学的研究者们通过使用常染色和X染色体、父系Y染色体、母系线粒体等等各类遗传标记体系来解析东亚人群的遗传多样性。
常染色体和X染色体遗传自父母双方,会被重组所打乱,而Y染色体上主干的非重组区呈严格父系遗传,并且Y染色体的“有效群体大小”理论上至多为常染色体的四分之一,X染色体的三分之一,对漂变非常敏感,容易形成群体特异性多态标记,从而包含更多的关于群体历史的信息。
Y染色体的这些特点使其成为研究人类进化和迁徙最强有力的工具之一。
Y染色体进入人们的视野,开始于其在追溯现代人起源上的应用。
自上世纪90年代以来,人类学界争论最激烈的话题,是东亚地区现代人的起源问题。
由于东亚出土了大量的古人类化石,一些人类学家认为东亚地区的人类是本土连续进化的,支持全球现代人的多地区起源。
然而,1999年宿兵等人采用Y染色体非重组区的19个SNP来研究东亚人群,得出东亚地区现代人起源于非洲,并由南方进入东亚,而后向北方迁。
随后,2000年柯越海等人的对东亚地区12127份男性随机样本的Y染色体进行SNP分型研究。
Y染色体突变M168被认为约是7.4万年前现代人走出非洲时所产生的突变,其原始型仅出现在东非人群中,除非洲以外的人群都是突变型。
柯越海等人的研究虽然没有直接检测M168这个突变,但他们检测了M89、M130和YAP这三个M168下游的突变。
从分子人类学发展史看人类文明的y

[转载]从分子人类学发展史看人类文明的Y1 分子人类学发展简明史(摘自网络资料整编)分子人类学产生于本世纪60 年代,它是分子生物学与人类学交叉产生的边缘学科。
从60 年代开始,一些分子生物学家逐步将分子生物学技术引入人类学研究领域,试图通过研究人类DNA 中所蕴藏的遣传信息来揭示整个人类的形成与演化过程,并寻找人类的祖先。
随着分子生物学理论和技术应用于人类学领域,人类学的一门新的分支学科——分子人类学Molecular anthropdogy )兴起了。
关键进展,三次大争论:初显威力,人猿分离时间大争论:分子人类学在诞生之初并未受到人类学界的欢迎。
许多人类学家对该学科的研究成果都持怀疑或否定态度。
60 年代未美国加州大学伯克利分校的两位分子生物学家一阿伦•威尔逊(Alan Wilson,后来夏娃理论的主要提出者)和文森特•萨里奇(Vincent Sarich )通过比较现代人类和非洲猿类的血液蛋白的分子结构,得出了人猿分离时间为距今约500 万年前的结论。
这与古人类学家通过化石证据获得的1500 万年前的结论相去甚远,因而引发了人类学界的一场大争论。
当时绝大多数古人类学家都不相信威尔逊和萨里奇的结论,认为这是离奇和荒谬的。
但在以后的十多年中,其他一些分子生物学家通过研究人类其他组织的分子结构,得出了和威尔逊等相似的结果。
80 年代初,匹尔比姆(Pilbeam )和安德鲁斯(Andrews )根据新的化石材料否定了腊玛古猿Ramapithecus, 当时曾认为其生存年代为1400 万年前)的人类祖先地位,从而大大缩短了我们人类出现的时间。
整个人类学界不得不承认:在人猿分离问题上,威尔逊和萨里奇是正确的,而人类学家是错误的。
此后,一些古人类学家(如英国的斯琼格Stringer 、德国的布劳尔Brauer 等)开始欢迎分子生物学家的介入,并把分子人类学的研究成果视为人类起源研究的重要组成部分。
夏娃理论,人类走出非洲:在人猿分离争论结束后不久,一场更大的争论席卷了整个人类学界,这场争论一直延续至今。
人类Y染色体研究进展

-8-
SRY ZFY MIC2 TP2
AMELY TPSY HY IL9R
GCY AZFa (USP9Y , DBY) AZFb(YRRM1) AZFd AZFc(DAZ)
-9-
-10-
Y染色体基因分类
1.pseudvaatosomal基因, 基因, 基因 如MIC2 ,IL9R… 2.位于 位于X-Y同源区的 同源区的NRY基因, 基因, 位于 同源区的 基因 如DBY, USP9Y ,UTY… 3.特有基因, 特有基因, 特有基因 如SRY基因 基因
-13-
Y染色体的自我保护 染色体的自我保护
Y染色体上的所有基因 染色体上的所有基因 都可能受到层出不穷的X 都可能受到层出不穷的 基因的攻击,最后Y染色 基因的攻击,最后 染色 体丢盔弃甲, 体丢盔弃甲,只得把绝大 部分已被瞄准的基因“ 部分已被瞄准的基因“关 闭”或者切除。这就是Y 或者切除。这就是 染色体越来越短的原因。 染色体越来越短的原因。
染色体结构异常病: 三. Y染色体结构异常病: 染色体结构异常病 • Y染色体的长臂或短臂缺失 染色体的长臂或短臂缺失 • 等臂染色体 (Yq)和i(Yp) 等臂染色体i( ) ( ) • 环状染色体 • 双着丝粒染色体 • 倒位和各种涉及 染色体的易位 倒位和各种涉及Y染色体的易位 • …
-25-
-14-
在人类基因组测序计划初期, 在人类基因组测序计划初期,科学家们发现 Y 染色体的中间区段有太多太多的重复基因 中间区段有太多太多的重复基因和没有 染色体的中间区段有太多太多的重复基因和没有 任何生理功能的惰性基因 后来发现Y染色体上 惰性基因, 任何生理功能的惰性基因,后来发现 染色体上 存在的这些基因实际上就是一些所谓的遗传密码 回文结构。例如ABCDEFG和GFEDCBA就构 的回文结构。例如 和 就构 成了一对回文结构,也可以说互成镜像。Y染色 成了一对回文结构,也可以说互成镜像。 染色 体就是依靠这些回文结构来保持自身遗传信息的 稳定性的。 稳定性的。
因纽特人y染色体单倍群dna的类型

因纽特人y染色体单倍群dna的类型标题:探寻因纽特人y染色体单倍群DNA的类型及其文化意义导言1. y染色体单倍群DNA的概念和重要性在人类遗传学研究中,y染色体单倍群DNA是指由父系传承的DNA序列,通过对不同群体的y染色体单倍群DNA进行分析,可以揭示人类个体之间的遗传联系,同时也能够为人类种群的起源、迁徙和遗传多样性提供重要线索。
因纽特人作为北极地区的土著民族,其y染色体单倍群DNA类型的研究对于揭示他们的遗传渊源、文化演变及人类学研究有着重要意义。
2. 因纽特人的历史和文化背景作为北极圈内的原住民族,因纽特人(Inuit)主要居住在加拿大、格陵兰和阿拉斯加等地,他们有着悠久的狩猎和捕鱼传统,过着以捕鱼、游牧和海洋产物为生的生活。
在长期的气候环境适应过程中,因纽特人形成了独特的文化和社会制度,传承着丰富的口头传统和宗教信仰,其社会结构和生活方式也受到地理环境和资源条件的影响。
3. 研究目的和意义本文将从因纽特人y染色体单倍群DNA的类型出发,探讨他们的遗传源流、族群关系及文化发展,通过对y染色体单倍群DNA的研究,寻找因纽特人文化与遗传的联系,进一步了解他们的起源和历史。
正文一、因纽特人y染色体单倍群DNA的类型介绍根据相关研究,因纽特人的y染色体单倍群DNA类型主要包括A2a、C2a、Q1a等,在这些类型中,A2a是最为常见的,这与他们亲缘关系的研究结果相一致。
对不同因纽特人群体的研究发现,这些y染色体单倍群DNA类型的分布存在一定的地域差异,在不同地区形成了特定的遗传分布格局。
二、因纽特人y染色体单倍群DNA与世界各地其他人群的比较通过与其他世界各地人群的y染色体单倍群DNA比较分析,可以发现一些有趣的现象。
比如与东亚、西伯利亚和北美印第安人的关系密切,同时又与北欧以及格陵兰因纽特人有着一定的联系等。
这些比较分析可以为因纽特人的迁徙史和遗传亲缘关系提供重要线索。
三、因纽特人y染色体单倍群DNA的文化意义在因纽特人的文化传统中,包括狩猎、图腾崇拜、宗教信仰等诸多方面都与他们的遗传联系密切。
人类染色体的传奇故事-可怜的Y染色体

人类染色体传奇故事:可怜的Y染色体在人类中,起到区分个体性别的主角就是性染色体:X和Y染色体。
人类是二倍体生物,其染色体都是成双成对存在的,每对染色体都是几乎没有形状差异的孪生姐妹。
但是X和Y这一对性染色体就显得有点离经叛道了:按照从大到小的染色体编号模式,在23对染色体成员中,X染色体应坐在第8把交椅上,但是与它搭档的Y染色体只有它大小的三分之一,甚至比最小的22号染色体还要小一点。
如果你是女性,性染色体是一对XX,那么一切正常,在女性细胞的复制与繁殖中,X染色体的行为和其它22对常染色体基本上没有差别:一样的两两互补配对,重组交换,你中有我,我中有你,彼此间朝着消弭差异的共同方向行事。
但是一旦Y染色体加入遗传信息的阵营,性染色体变成一对XY,一切就改变了。
Y染色体的个子太矮,根本无法与它的搭档X染色体进行完全的配对与重组互换,得委屈X染色体弓下腰来弯成一个圆环,然后蜻蜓点水一般与小Y染色体在顶端少量地配对、重组互换一下。
这样的交换对于维护Y染色体长久的稳定性是不够的,因为在染色体的遗传规律中,如果没有重组互换的行为发生,就意味着走上灭亡的道路。
那么Y染色体是如何保持长久稳定性的?这个秘密直到2003年才被科学家揭开,原来Y染色体的独特之处在于能够自身进行基因的重组互换。
除了模样与行为上的不同以外,在携带的基因种类与数量上也有巨大差异。
如Y染色体携带有启动男性形成发育的关键基因:SRY基因,在X染色体上没有这样的基因。
X染色体能够携带2000到3000个基因,可怜的Y染色体只能携带20到30个基因。
并且,Y染色体上的核苷酸序列看起来根本就是一堆毫无意义的垃圾山,很难找到基因的宝藏。
这样的特性一度令人类基因组测序计划阻滞不前。
性染色体在女性和男性细胞中具有这样巨大的差异,会不会因此造成遗传的紊乱呢?生命从来就是一个高度平衡体,女性细胞为了维持X染色体上的基因剂量与男性的一致,在普通的生活细胞中选择其中一条X染色体蜷缩起来,就像将不用的衣服装进衣橱,并挂上一把锁,封存。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
关于人体Y染色体的进化历程的研究报告研究课题:人类Y染色体的进化历程研究成员:文章撰写人:人类属于XY性别决定类型,在正常男性体内X与Y染色体是23对同源染色体中形状差别最大的一组。
人体内的Y染色体在形状上比X染色短。
单从数据上看,X在23对染色体中要排上第8位,但与其配对的Y染色体却只有它的三分之一,比最小的22号染色体还要小。
Y是惟一一个对个体生存不必要的染色体。
人如果没有X,或者没有其他任何一对常染色体,就无法出生,在胚胎期就会死亡。
而没有Y染色体却对个体生存没有关系,比如女性都没有Y。
其他染色体一般互相依靠,但是它们不依靠Y染色体。
2005年,X染色体基因草图显示,Y染色体上有用的基因部分转移到了X染色体上。
Y染色体甚至被戏称为X染色体的“错误版本”。
实际上,现在Y染色体上最有用的基因仅剩SRY基因(决定了睾丸的发生)此外,Y染色体还有抗癌作用,那么Y染色体的起源和发生是怎么样的呢?我们知道无脊椎动物是没有性染色体或是没有雌雄异体的机制,包括许多属于变温动物的脊椎动物也是没有性染色体的。
这些动物的性别许多不是受到基因型的控制的,他们的性别决定是受到环境影响的:像是一些动物的性别决定取决于温度——海龟的性别决定依赖于孵卵时的温度;对于盐生钩虾来说,在5℃下后代中雄性为雌性的5倍,而在23℃以下的后代中雌性是雄性的13倍。
故在生物的进化历程中性染色体的出现,是进化史上的一项重大事件。
XY染色体决定机制是高等生物标志(相比于其他的性别决定机制,如:ZW,XO等),可见Y染色体在生物进化上的重要地位。
根据《自然》杂志上一篇关于Y染色体进化的研究报告显示:Y 染色体的进化速度比其他的基因快2%,科学家通过将人类的基因与最像人类的“近亲”猩猩相比对,发现Y染色体与猩猩的差异度为30%,而人类其它基因与猩猩的差异度为28%,这也就是说人类的Y 染色体与人类其它基因相比存在2个百分点的进化程度差异。
而这一进化过程是从大约600万年前开始的,其他的基因的进化时间要比Y 染色体长的多。
研究报告的撰写者,剑桥大学怀特黑德研究所主任戴维·佩奇和麻省理工学院的一名教授表示:“看起来Y染色体是人类染色体中进化最快的一个,它几乎就是一个不断变化的基因,就像是一座不断重建的房屋一样。
动物的雄性祖先最早可以追溯到5亿年前,而Y染色体的出现不到3.1亿年,目前创造雄性所必须的基因出现可能不到1.7亿年。
3亿年前的某一天,在其中一条原始性染色体上一个叫SOX3的基因发生突变,变成了名为SRY的基因。
SRY基因是现代Y染色体上决定男性性别的关键基因。
然而最新的研究表明XY的决定系统大约在1.66亿年前出现。
科学家们利用动物分类学推测了基因的发展。
爬行动物的性别不是由性染色体决定的,它们通常由环境决定。
例如乌龟、鳄鱼等,在阳光充足的水滩上发育成雄性;在阴凉的背阴环境下发育成雌性。
比爬行类高级一点的是哺乳动物中的单孔类动物,如著名的鸭嘴兽、针鼹等。
单孔类动物是具有Y染色体的最古老动物,它们的性别不再是简单地由环境决定。
而哺乳动物与爬行类动物的分支进化发生在3亿年前,所以科学家们推断出Y染色体出现在那个年代。
科学家们也尝试了利用植物来进行研究:“人类的Y染色体在经历了大概200万年的进化史后,失去了大量的基因,现在只保留约3%的其祖先的基因。
然而,我们对早期阶段的过程知之甚少,因为它发生在很久很久以前,多伦多大学生态学与进化生物学系(EEB)的斯宾塞巴雷特教授说。
“包括人类和其他动物物种的Y染色体研究发现,退化始于数亿年前。
但植物并不是这样。
”具有独立性别的植物出现是一个相对较新的进化创新,这使它们非常适合这项研究。
巴雷特说。
“世界上大约只有百分之六的开花植物具有雄性和雌性之分,其余都是雌雄同体。
”科学家使用一种植物,它们大约15万年前左右首先进化出由X 和Y染色体控制的独立性别系统,相对于那些动物,它们显得年轻一些。
“我们测试Rumex hastatulus的Y染色体退化,Rumex hastatulus是来自美国南部的一年生植物,俗称heartwing sorrel。
我们发现,位于Y染色体的基因已经开始经历遗传性的退化,尽管它们的起源日期相对较新。
”这项研究的主要作者、多伦多大学生态学和进化生物学的博士生乔希霍夫说。
“重要的是,我们的研究结果表明,这种退化的程度取决于位于性染色体的基因停止相互重组的时间。
”之后科学家们再应用分子生物学上的分子进化钟来估计,发现X 和Y染色体上的SOX3基因与SRY基因也有了3亿年的分家史。
SOX3的基因突变成了SRY的基因后,又发生了一个染色体倒转事件,使本是同根生的SOX3基因(在X染色体的底部)与SRY基因(在Y染色体的顶部),河汉相隔,老死不相往来。
染色体倒转是染色体内部区段发生重组的一种较常见方式,这种行为常常导致染色体上基因的位置发生重新排列。
而在Y染色体上发生的倒转基因又不像X染色体一样还可以在雌性的细胞中得到孪生姐妹的修复帮助,所以这些倒转了的区段还常常被Y染色体自身给切除掉了,结果随着时间的流逝,在没有备份可以弥补的情况下,可怜的Y染色体失掉了越来越多的基因,变得越来越萎缩,到今天只有X染色体的三分之一。
在我们的鸭嘴兽祖先出现了Y染色体后,这条执意走独木桥的Y 染色体就常常发生各种各样的基因倒转和删除事件,使得Y染色体上的基因越来越与众不同。
在人类中的雄性中,Y染色体上只有两头的部分还保留着与X染色体进行重组互换的能力,而雌性的两条基因分布均一的X染色体在所有的部位仍进行着充分的重组互换。
X染色体和Y染色体之间的基因重组已被证实是对生命体有害的,它会导致雄性动物丢失Y染色体在重组之前所含有的必需基因、雌性动物多出原本只会出现在Y染色体上的非必需基因甚至是有害基因。
所以,在进化过程中,对雄性有利的基因就逐渐在性别决定基因附近聚集,后来这个区域的基因发展出了重组抑制机制以保护这个雄性特有的区域。
Y染色不断体沿着这种路线演化,抑制Y染色体上的基因与X染色体上的基因发生重组。
这个过程最终使得Y染色体上约95%的基因不能发生重组。
同源染色体的基因重组本是用于降低有害突变保留的几率、维持遗传完整性的,但Y染色体因不能与X染色体发生重组,被认为容易发生损毁而导致退化。
人类的Y染色体在其演变的过程中丢失了原本拥有的1,438个基因中的1,393个,约每一百万年丢失4.6个基因。
据推算,若Y染色体仍以这样的速率丢失基因,它有可能在一千万年后完全丧失功能。
我们以创造精子的UBE1基因为例,几乎所有的哺乳动物都通过这种基因制造精子。
然而科学家们发现我们人类以及黑猩猩和其他灵长目动物都缺乏UBE1基因。
科学研究者认为,大约在四千万年前,我们的灵长目祖先丢失了UBE1基因,它的工作被另一个染色体的另一个基因代替。
另一方面,男性Y染色体上的基因丢失也并非只起负面作用,它实际上是在优化基因。
例如,Y染色体上有一种为耳毛编码的基因,有了这种基因,才会长出耳毛。
而有些男人可能还会有耳毛,但大部分男人已经没有耳毛了,因为耳毛完全是多余的。
Y染色体上丢失的都是这些对人没有太大作用的基因。
Y染色体的确虽然是人类所有染色体中最为短小的一个,但它的基因结构并未完全为人所知。
在人类基因组测序计划初期,科学家们发现,Y染色体的中间区段有太多太多的重复基因和没有任何生理功能的惰性基因,要想从片区域鉴定出某个具体基因,实在是一件难事。
一位致力于Y染色体研究的科学家形象地说,分析Y染色体上的基因,就像让你走入一个由镜子搭起来的小屋里,到处都是一个相同而成镜像的影子,甚至无法辨清天花板和地面。
直到2003年,科学家才搞清,Y染色体上大量的“影子”基因实际上就是一些所谓的遗传密码的回文结构。
例如ABCDEFG和GFEDCBA就构成了一对回文结构,也可以说互成镜像。
这些让人无法辨别的“影子”基因,在保证弱小的Y染色体亿万年不再继续退化做出了巨大贡献。
其它所有染色体都具有与其孪生伙伴配对重组互换的能力,Y染色体则只能在两端与它的伙伴X染色体重组互换而得到修复与保存。
依靠这些回文结构,小小的Y染色体可以从中间弯起来,让自身对等回文部位配对并发生重组互换,如果中间某个基因不幸突变,他就可以从它对面的回文备份中得到修复。
进化之路常常是迅速而复杂的,科学家无法做出准确的推测,然而未来Y染色体的命运确是显而易见,自Y染色体诞生以来,每过一百万年,Y染色体上就有三到六个基因灭失掉,按照这个速度进行下去,Y染色体只剩下一千万年的寿命了。
然而,Y染色体的死亡并不意味着男性的终结。
我们只要看一下科学家们对于鼠类的研究便可知:几种同属的鼠科及仓鼠科的啮齿目动物已经通过下列途径达到Y染色体演化终端:土黄鼹形田鼠(Ellobius lutescens)、坦氏鼹形田鼠(Ellobius tancrei)及日本刺鼠中的奄美刺鼠(Tokudaia osimensis)和冲縄刺鼠(Tokudaia muenninki),已完全丢失它们的Y染色体(包括SRY基因)。
裔鼠属(Tokudaia)下的一些鼠类将其余的一些原来在Y染色体上的基因转移到了X染色体上。
土黄鼹形田鼠和裔鼠属的鼠类中不论雄性或雌性的基因型皆为XO,而所有坦氏鼹形田鼠的基因型皆为XX。
这些啮齿目动物的性别决定系统仍未被人们完全了解。
林旅鼠(Myopus schisticolor)、鄂毕环颈旅鼠(Dicrostonyx torquatus),和南美原鼠属(Akodon)中的众多物种通过X染色体和Y染色体复杂改变,演化出除了基因型为XX的雌性以外的另一种拥有一般雄性才拥有的XY基因型的雌性。
在雌性潜田鼠(Microtus oregoni)中,每个个体的单个体细胞只有一条X染色体,只产生一种X配子;而雄性的潜田鼠基因型仍为XY,但可以通过不分离现象(Nondisjunction)产生Y配子和不含任何性染色体的配子。
无人知道鼠类是怎样在进化过程中成为第一类没有Y染色体的哺乳动物,但有一点我们几乎是可以肯定的:鼠类在其他的染色体上已经进化出了负责制造雄性的新基因,其中就有一种新基因接替了SRY基因的工作。
在啮齿目动物之外,黑麂(Muntiacus crinifrons)通过融合原有的性染色体和常染色体演化出了新的X染色体和Y染色体。
灵长目动物(包括人类)的Y染色体已严重退化这一现象预示着,这类动物会相对较快地发展出新的性别决定系统。
学者估计,人类将在约1.4千万年后获得新的性别决定系统。
“如果人类能够度过Y染色体灭亡带来的灾难,那也将有极大的变化,即种属变化。
也就是说,那以后的人,和我们现在的人不是同一种属了。
”北大生物学家饶毅这么断言。
为什么会有这样的说法呢?假如我们人类还能再存活一千万年的话,我们后代即使没有Y染色体,他们也会进化出制造男性的新基因,但这种进化之路并不可能是平坦的。