脊椎动物血液循环系统的演化
描述蛙的心脏和血液循环路线的特点。

主题:蛙的心脏和血液循环路线的特点一、蛙的心脏结构蛙的心脏是由三个部分组成的,分别是两个心房和一个心室。
两个心房位于心脏的上部,而心室则位于心脏的下部。
心房和心室之间分别由两个瓣膜相连,这些瓣膜可以控制血液的流动方向,确保血液在心脏中流动的顺畅性。
二、蛙的血液循环路线1. 肺循环蛙的血液循环路线是通过两个独立的循环系统完成的。
首先是肺循环,它起始于静脉回流的血液进入右心房,然后通过右心房收缩将血液送入右心室,接着右心室的收缩将血液推入肺动脉。
血液随后通过肺动脉进入肺部进行气体交换,在此过程中二氧化碳被释放掉,氧气则被血液吸收。
氧合的血液随后通过肺静脉进入左心房,最终进入左心室。
2. 体循环体循环起始于左心室的收缩,将氧合的血液通过主动脉送入体内各个组织器官。
在组织器官中,氧合的血液释放氧气,同时吸收二氧化碳。
经过这样的气体交换后,血液变成含有二氧化碳的静脉血,随后通过静脉进入右心房,重新开始新一轮的循环。
三、蛙的血液循环路线的特点1. 双循环系统蛙的血液循环路线采用双循环系统,即肺循环和体循环分开进行。
这种双循环系统有助于将氧合的血液分别送到肺部和体内各个器官,从而有效地实现气体交换和供给氧气的功能。
2. 三腔式心脏蛙的心脏结构较为特殊,采用了三腔式结构,即两个心房和一个心室。
这种心脏结构使得氧合的血液和含有二氧化碳的血液在心脏中可以得到有效的隔离,减少了混合的可能性,有利于维持血液的氧合程度。
3. 顺应性蛙的心脏和血液循环系统具有较高的顺应性,即在不同的情况下能够根据需要调整心脏的收缩力度和血液的流动速度,以满足身体对氧气和营养的需求。
这种顺应性有利于适应不同环境条件下的生存。
蛙的心脏和血液循环系统具有双循环系统、三腔式心脏和顺应性等特点,这些特点使得蛙能够有效地维持身体的氧气供应和二氧化碳的排出,从而适应各种生存环境。
蛙的心脏和血液循环路线的特点所蕴含的生物学奥秘蛙作为两栖类动物,其心脏和血液循环系统的特点不仅仅是一种生理结构的设计,更是适应其生存环境和生活习性的生物学奥秘。
4第十章动物的循环、呼吸和排泄系统

1.3.2 人心脏的结构
心脏是十分重要的器官,是循环系统的 总枢纽,其收缩和舒张造成血液的循环流 动。心博一旦停止,血液循环不能进行, 各处组织不能获得营养,也不能排除废物, 生命就很快完结。 人的心脏重约400克左右。位于胸腔的 围心腔中。 心脏分为四室,即左心房左心室和右心 房右心室。左右两半界限分明。右心房右 心室的血液是从大静脉流入的带二氧化碳 的血,左心房左心室的血液是从肺流回的 带氧的血。 但它们的博动却是心房与心室分别同步。 即左右心房先同时收缩,然后左右心室再 同时收缩。
心肌的特殊传导系统 数字表示兴奋从窦房结传 递到该点的时间(秒)
Ø 心肌细胞的动作电位
以浦肯野氏细胞为例: 0期——细胞快速去极化(动作 电位的升支) 1期——短暂而快速的复极化 2期——缓慢复极化(平台期) 3期——较快复极化 4期——复极化至静息膜电位 浦肯野氏细胞的动作电位
自律细胞到达静息膜电位水平 后又开始缓慢去极化,直到阈电 位水平,再引起心肌细胞的兴奋, 又开始出现新的动作电位。
Ø Rh因子
Rh是恒河猴(Rhesus Macacus)外文名称的头两个字⺟。兰德斯 坦纳等科学家在1940年做动物实验时,发现恒河猴和多数人体内的红 细胞上存在另一种抗原物质,命名为Rh因子。 凡是人体血液红细胞上有Rh因子者,为RH阳性。反之为RH阴性。 RH阳性血型在我国汉族及大多数民族人中约占99.7%,个别少数民族 约为90%。在国外的一些民族中,RH阳性血型的人约为85%。
Ø 血液成分
淡⻩色液体,约占血液体积的53%,其中水分 血浆(plasma) 约占92%,其余8%为溶于水的晶体物质(无 机盐)和胶体物质等(血浆蛋白)。 颗粒细胞(嗜中性、嗜酸性、嗜碱性粒细胞) 白细胞 无颗粒细胞 淋巴细胞 起防御作用
生物学习题及答案

第一部分:名词解释生物膜;干扰素;稳态;光周期诱导;光合磷酸化;光敏色素;无氧呼吸;细胞呼吸;菌根;双受精;生物节律;等位基因;细胞分化;基因库;非共质体途径;内皮层;无氧呼吸;再生作用;适应;原核细胞;氧化磷酸化;底物水平的磷酸化;体液免疫;形成层;克隆;共质体途径;细胞周期;三羧酸循环;世代交替;蛰伏;基因库;内皮层;无氧呼吸;再生作用;适应;应激性;蛋白质的一级结构原肠胚;中心法则;内起源;协同进化;成花素;光能细菌;病毒粒子;反馈调节;基因突变;细胞外消化;蛋白质的二级结构;光呼吸;春化作用;化能细菌;内吞作用;无限维管束;细胞分化;稳态基因文库;菌根;生态位;光系统;食物链;生物多样性;环境容量;群落;二次污染物;不可再生资源;种群;质壁分离;年轮;抗原;体循环;光合作用;光反应;暗反应;领地;行为;细胞克隆选择学说;达尔文自然选择学说;压力流假说;团聚体学说;内聚力学说;灾变论;大气圈;学习;血液循环;周围神经系统;腐食性营养;染色体组型;细胞骨架;酶;细胞周期;减数分裂;肺活量;有丝分裂;变态;生态金字塔;遗传漂变;基因工程;生物节律;微球体学说;本体感受器;生物钟;多倍体;拟态;渐变式进化和跳跃式进化;自然发生说;自然分类;五界系统;病毒和反病毒;原核生物和真核生物;原口动物;后口动物;生态系统;生态幅;最低量定律;寄生和共栖;化学互助和拮抗;生态位顶级群落;生物地化循环;稳态;耗散结构;生物大分子;胞饮作用端粒;内环境;细胞内消化和细胞外消化;干细胞;反射弧;光周期双受精;孤雌生殖;孢原细胞;缺失;重复;倒位;易位;上位效应;抑制基因;互补基因;转化;中心法则;操纵基因;结构基因;遗传漂变;异地物种形成;协同进化;趋同进化;共进化;趋异进化;人口问题第二部分:填空题⒈细胞呼吸全过程可分为糖酵解、、和电子传递链。
⒉细胞核包括核被膜、、和核仁等部分。
⒊消化系统由消化管和两部分组成。
动物学思考题级答案

动物学思考题级答案动物学思考题⼀、动物的分类和系统发⽣1.物种(species)2.⼆名法,举例说明3.简述⽣物界的5界系统4.简述动物界的主要类群⼆、动物体的基本结构1.简述动物体的基本结构机制及其发展进化趋势动物的对称类型可分为;动物的体腔类型可分为;同律分节、异律分节、⾝体分部。
2.真体腔动物真体腔(Eucoelomata);裂体腔法和肠腔法;原⼝动物与后⼝动物。
重要名词:物种,⼆名法,五界系统,侧⽣动物,分类阶元,分类⽅法,系统发⽣,个体发⽣3. 多细胞动物的胚胎发育3.1 动物的完全卵裂的2种主要形式和特点动物的完全卵裂有两种主要模式:即辐射卵裂和螺旋式卵裂。
卵裂⽅式的不同,往往会影响到胚胎后期的发育。
3.2 动物的早期胚胎发育的⼏个主要阶段及发育过程的特点囊胚(blastula):囊胚层(blastoderm),囊胚腔(blastocoel),原肠胚(gastrula):原肠腔(gastrocoel),中胚层和体腔(mesoderm and coelom) ,神经胚(neurula)。
3.3 动物的中胚层的发⽣中胚层的形成⽅式主要有2种:(1)体腔囊法(2)裂体腔法3.4 假体腔与真体腔3.5 简述神经胚的形成过程4.试⽐较原⼝动物与后⼝动物的差别胚孔(blastopore),原⼝动物(Protostomia)和后⼝动物(Deuterostomia)。
三、动物的系统与进化(各门类的特征与进化关系)1. 原⽣动物门2.海绵动物门3.腔肠动物门4.扁形动物门5. 原体腔动物6.环节动物门7.软体动物门8.节肢动物门9.棘⽪动物四、各门动物的分述1. 原⽣动物门(Protozoa)1.1原⽣动物的主要⽣物学特征及其在⽣物系统进化中的地位1.2 区别纤⽑⾍纲、鞭⽑⾍纲、⾁⾜⾍纲原⽣动物要点特征1.3 简述纤⽑和鞭⽑的结构及其在原⽣动物⽣活中的作⽤1.4 原⽣动物的⽔分调节和排泄1.5 纤⽑⾍的接合⽣殖和⽆性⽣殖的过程1.6 原⽣动物的⽆性⽣殖⽅式1.7 昏睡病、⿊热病是哪⼀类原⽣动物引起的疾病?(我国⼈体五⼤寄⽣⾍)1.8 原⽣动物分布在那些环境中1.9原⽣动物在污⽔处理系统中所起的作⽤1.10 简述疟原⾍的⽣活史,引起⼈类发病原因及症状(1)红细胞前期(2)红细胞外期(3)红细胞内期引起⼈发病的原因及症状:1.11 哪类原⽣物中的种类有外壳1.12 孢⼦⾍纲与丝孢⼦⾍纲的区别1.13 纤⽑⾍感受外界刺激和防御的结构2. 侧⽣动物—海绵动物门(Spongia)2.1 为什么说海绵动物是多细胞动物进化中的⼀个侧枝?2.2 描述海⾯动物的体壁结构2.3 海绵动物⽔沟系的结构和功能2.4 海绵动物⽔沟系是如何进化的?2.5 以钙质海绵为例,说明海绵动物早期胚胎发育的过程2.6 海绵动物分为哪纲及主要区别2.7 海绵动物的体制与其⽣活⽅式的适应关系3. 腔肠动物门3.1 ⽐较腔肠动物中⽔螅型和⽔母型的异同3.2 为什么说腔肠动物的细胞出现了初步的组织分化?3.3 举例说明世代交替和多态现象3.4 如何区分⽔螅纲⽔母和钵⽔母纲⽔母?3.6 如何确定腔肠动物的进化地位?3.7 简述腔肠动物的主要特征3.8 如何看待栉⽔母动物的进化地位4. 三胚层⽆体腔动物4.1 扁形动物门(Platyhelminthes)的主要特征4.2 ⽪肤肌⾁囊的结构4.3 形动物的中胚层分化的组织和器官4.4 扁形动物神经系统的特点4.5 原肾管型排泄系统的结构特点4.6 扁形动物门分纲及各纲的主要特征4.7 扁形动物⾼度适应寄⽣⽣活的特征4.8 寄⽣⾍和寄主之间的相互关系及防⽌原则4.9 为什么说三胚层⽆体腔动物是动物系统进化中的⼀个新阶段重要名词:原肾管,杆状体,囊尾幼⾍,厌养呼吸,中间寄主,终末寄主,幼体⽣殖,假分节,不完全消化管,牟勒⽒幼⾍。
脊椎动物五大类的心脏的结构和血液循环途径
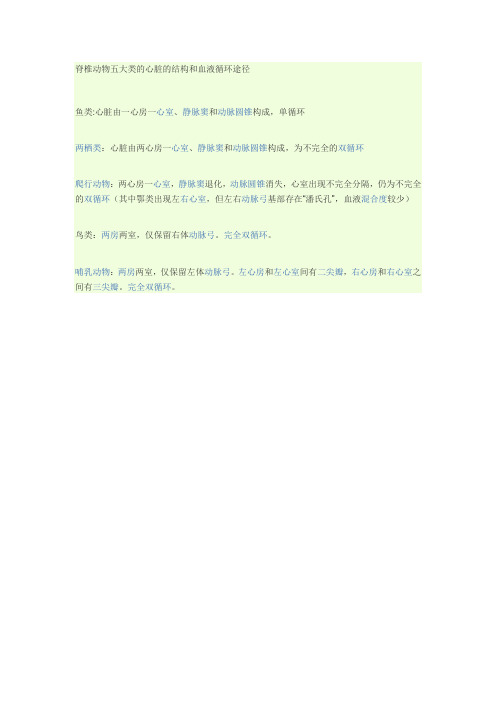
脊椎动物五大类的心脏的结构和血液循环途径
鱼类:心脏由一心房一心室、静脉窦和动脉圆锥构成,单循环
两栖类:心脏由两心房一心室、静脉窦和动脉圆锥构成,为不完全的双循环
爬行动物:两心房一心室,静脉窦退化,动脉圆锥消失,心室出现不完全分隔,仍为不完全的双循环(其中鄂类出现左右心室,但左右动脉弓基部存在“潘氏孔”,血液混合度较少)
鸟类:两房两室,仅保留右体动脉弓。
完全双循环。
哺乳动物:两房两室,仅保留左体动脉弓。
左心房和左心室间有二尖瓣,右心房和右心室之间有三尖瓣。
完全双循环。
脊椎动物血液循环系统的演化

脊椎动物血液循环系统的演化一:心脏⒈位置:心脏位于体腔前部,消化管腹侧的一个围心腔中,由围心膜所包被。
鱼类和有尾两栖类的围心腔位于体腔前方。
陆生脊椎动物的心脏向后,向腹方移动至体腔的前腹位。
⒉结构:鱼类的心脏由静脉窦,一心房,一心室,动脉圆锥组成。
两栖类演变为两心房一心室,心房内出现完全或不完全的房间隔,静脉窦和动脉圆锥仍存在。
爬行类的心脏包括完全分隔的2个心房1个心室和退化的静脉窦,动脉圆锥消失,心室出现不完全分隔。
鸟类和哺乳类的心脏完全的分为四室,即左右心房和左右心室,其中哺乳类的左右心室之间有二尖瓣,左右心房之间有三尖瓣。
二:血液循环鱼类的血液循环为单循环,即由心室压出的缺氧血经入鳃动脉进入鳃部进行气体交换,出鳃的多氧血不再回心脏而是经出鳃动脉直接沿背大动脉流到全身,从各组织器返回的缺氧学经主静脉系统再流回心脏,形成一个大圈。
两栖类为不完全双循环,左心房接受从肺静脉返回的多氧血,右心房接受从体静脉返回的缺氧血以及皮静脉返回的多氧血,它们最后均进入心室。
爬行类仍为不完全双循环。
鸟类和哺乳类为完全双循环,从体静脉回心的缺氧血经右心房进入右心室,被压入肺动脉弓,从肺静脉回心的多氧血经左心房进入左心室,被压入体动脉弓。
三:动脉系统动脉系统的基本模式:腹大动脉,背大动脉,动脉弓胚胎期一般为六对动脉弓的演变:鱼类:由于以鳃呼吸,动脉弓在鳃部断裂为两部分,即入鳃动脉和出鳃动脉,中间以毛细血管联系,以进行气体交换。
软骨鱼类保留第2至6对动脉弓,硬骨鱼类保留第3至6 对动脉弓,其余退化。
两栖类以上的脊椎动物:成体因营肺呼吸,动脉弓不再断开,并只保留第3,4,6对动脉弓。
第三对为颈动脉,分布于头部和脑;第四对为体动脉弓,左右体动脉汇合成背大动脉;第六队为肺动脉弓;其中鸟类成体仅保留右体动脉弓,哺乳类则保留左体动脉弓。
四:静脉系统⒈鱼类:具H型主静脉系统,一对前主静脉窦,一对后主静脉窦,一对总主静脉,最后汇入静脉窦。
脊椎动物各系统演化
腰带组成骨盆。肩带中的肩胛骨更为发达。锁骨变化多。肢骨的基本情况未变,唯腕骨数目减少。
二、鱼类、两栖类、爬行类、鸟类和哺乳类的消化系统
观察液浸标本,Biblioteka 较五类动物消化器官的口裂和口腔、消化管的各部分及消化腺。
三、鱼类,两栖类,爬行类,鸟类和哺乳类的呼吸系统(图5—19)鱼类:
呼吸器官为鳃,受鳃弓和鳃条支持,鳃前隔的两面具有许多行平行褶皱的鳃瓣。内中有很多微血管,颜色鲜红,是气体交换的场所。
3)呼吸道和消化道逐渐趋于分开:
①两栖类的呼吸通道和食物通道在口咽腔处形成交叉。
②爬行类的鳄到哺乳类,形成了次生颚,内鼻孔后移,呼吸道和消化道完全分开。
4)呼吸道进一步分化,发声器逐渐完善。
脊椎动物心脏各部分的比较:
圆口类
静脉窦
有有有退化并入心房
动脉圆锥无有动脉球
心房111222
心室111(有室间隔)2
脊椎动物各系统演化
一、鱼类,两栖类,爬行类、鸟类和哺乳类的骨骼
观察经制备好的骨骼标本,了解其特点。
1.主轴骨骼
鱼类:
脊柱分躯椎(附有肋骨,保护内脏器官)和尾椎(运动用)两部。
两栖类;脊柱分?化为一块颈椎、七块躯椎和——块骶椎,尾椎则愈合为一块尾杆骨。
爬行类:
脊柱分化为颈椎、胸腰椎、骶椎及尾椎。
鸟类:
4.鸟类:
大脑半球较前更发达,嗅叶退化,中脑和小脑也相当发达。
5.哺乳类:
大脑半球很发达,已形成大脑皮质,为最高神经中枢;中脑为四叠体;小脑很发达,侧叶表面上有皱褶出现。
1、颌弓与脑颅的连接方式:
双接式:
颌弓通过它本身和舌颌软骨与脑颅连接起来,见于原始的软骨鱼、总鳍鱼。舌接式:
动物学问答题
动物学(乙)问答题1.原生动物有何主要特征?为什么它们是最原始、最低等的动物?2.试比较原生动物门各纲的运动器官。
3.试述腔肠动物的主要特征。
4.试述扁形动物的主要特征。
5.中胚层和两侧对称出现在动物演化上有何重要意义?6.试述中胚层形成的意义7.试述疟原虫的生活史。
8.羊肝蛭的生活史。
9.猪带绦虫的生活史。
10.十二指肠钩虫的生活史。
11.比较实验中观察到蛔虫和蚯蚓体壁横切面上的主要不同点。
12.吸虫纲主要特征13.假体腔动物的基本概念和特征是什么?14.假体腔与真体腔有何不同?15.何谓次生体腔?它有何意义?16.线虫动物门的简要特征是什么?17.环节动物门的主要特征。
18.试述软体动物门的主要特征。
19.河蚌适于埋栖生活的特点20.试述节肢动物门的主要特征。
21.对节肢动物现有的6个纲作一分析比较22.以对虾为例说明甲壳纲的主要特征23.举例说明昆虫的生殖和变态24.试述无脊椎动物神经系统的演化趋势25.简述棘皮动物门的主要特征。
26.简述半索动物主要特点和半索动物在动物系统上的地位。
27.脊索动物门的共同特征是什么?28.试述脊椎动物亚门的主要特征。
29.试述鱼类主要特征和鱼类适应水生生活的特征。
30.鱼类的亚纲分类情况。
31.软骨鱼和硬骨鱼的特征32.简述侧线器官的作用33.试述两栖纲的主要特征。
34.试述爬行纲的主要特征。
35.羊膜卵的结构及羊膜卵的生物学意义是什么?36.为什么说爬行动物的循环系统也是不完善的双循环?37.试述鸟纲的主要特征。
如何适应飞翔生活?38.今鸟亚纲的有哪三个总目?特征如何?39.鸟类双重呼吸特点40.试述哺乳纲的主要特征。
41.哺乳动物三个亚纲的主要特征。
42.哺乳动物可分为哪三个亚纲,及其主要特征?并举代表种43.简述比较脊椎动物各系统:循环、呼吸、排泄44.比较脊椎动物呼吸系统的进化?45.比较脊椎动物循环系统的进化?46.鸟类的形态结构如何与其飞翔生活相适应的47.为什么说从爬行动物开始,才算是真正的陆生动物?动物学(乙)问答题1.原生动物有何主要特征?为什么它们是最原始、最低等的动物?从单细胞动物(细胞器)、分布广、营养(自养、异养)、呼吸、排泄、应激性和运动、生殖、包囊等方面阐述。
脊椎动物从水生到陆生的演化
一、呼吸的进化1、呼吸方式脊椎动物的呼吸方式可分为两大类,即水栖种类用鳃呼吸,在水与鳃上毛细血管内的血液间进行气体交换。
鳃分内鳃及外鳃二种类型,内鳃在园口类,鱼类终生存在,外鳃存在于所有两栖类的幼体及部分有尾两栖类的成体。
陆生种类用肺呼吸,在空气与肺上毛细血管内的血液间进行气体交换。
此外一些种类尚有辅助呼吸器官,如蛙的皮肤,乌鳢的口壁粘膜,泥鳅的消化管等。
2、呼吸特点a、鱼类:鱼类的鳃位于咽部两侧,由鳃弓支持着,每一鳃弓上有两列鳃丝(软骨鱼类第五对鳃弓只有1个鳃瓣,硬骨鱼类第五对鳃弓多特化为咽骨,其上无鳃丝)。
软骨鱼有鳃间隔自鳃弓伸到体表下,鳃瓣(软骨鱼的鳃瓣多不为丝状)附在鳃间两侧。
硬骨鱼鳃间隔退化,鳃丝附于鳃弓上,鳃裂被鳃盖骨所覆盖,以鳃孔通于体外。
水流从口进入以后流经鳃,水中的氧和血液中的二氧化碳进行交换。
氧进入血液中,而二氧化碳则随水流排出体外。
b、两栖类:幼体用腮呼吸,变态后,内腮消失,用肺呼吸。
鳃是由外胚层发育来的,而肺则是由原肠管突出的盲囊形成的。
因此,鳃与肺不是同源器官(同源器官是指起源相同,构造和部位相似而形态和功能不同的器官),而是同功器官(形态和功能相似,起源和构造不同的器官)。
鳔和肺才是同源器官。
虽然有些鱼类的鳔已执行肺的功能,但专门作为呼吸器官的肺则是起源于两栖类。
两栖动物的肺构造简单,仅为1对薄壁的囊(如蝾螈)或囊内稍有些隔膜(如蟾蜍)而已。
其表面积比较小,不足以满足两栖类对氧的需求。
因此,两栖类还需借助于皮肤呼吸来摄取更多的氧。
c、爬行类:爬行类的肺较两栖类进步,肺的内表面积相对比较大,这是由于肺内具有很多发达的隔膜。
一些结构高等的爬行类(如鳄和某些蜥蜴),肺内腔一再分割,腔内壁呈蜂巢状小室,从而扩大了与空气的接触面积。
由于开始形成了胸廓,靠肋间肌的收缩,胸廓的扩张与缩小,改变容积,从而使气体吸入或排出。
d、鸟类:鸟类的肺极为特殊,外观上看是一对海绵状体,内部则是由大大小小的各级支气管形成的彼此吻合相通的密网状管道系统和血管系统组成,称为网状管道肺。
动物循环系统
一、循环系统概念:循环系统(Circulatory system)是生物体的细胞外液(包括血浆、淋巴和组织液)及其借以循环流动的管道组成的系统。
循环系统是进行血液循环的动力和管道系统,由心血管系统和淋巴系统组成。
从动物形成心脏以后循环系统分心脏和血管两大部分,叫做心血管系统。
淋巴系统包括淋巴管和淋巴器官,是血液循环的支流,协助静脉运回体液入循环系统,属循环系的辅助部分。
二、动物循环系统发展历程从环节动物门开始出现,环节动物有次生体腔的出现,相应的促进了循环系统的发生。
环节动物具有较完善的循环系统,结构复杂,由纵行血管和环行血管及其分支血管组成,各血管以微血管往相连,血液始终在血管内流动,不流入组织间的空隙中,构成了闭管式循环系统。
血液循环有一定方向,流速较恒定,提高了运输营养物质及携氧机能。
软体动物门的循环系统由心脏、血管、血窦及血液组成血液自心室经动脉,进入身体各部分,后汇入血窦,由静脉回到心耳,故软体动物为开管式循环。
节肢动物门循环系统开管式,包括心脏和动脉两部分。
鱼的循环系统包括液体和管道两部分,液体是指血液和淋巴液,管道为血管及淋巴管。
两栖类由单循环的血液循环方式发展为包括肺循环和体循环的双循环,循环系统包括血管系统和淋巴系统两部分。
鸟类的循环系统反映了较高的代谢水平,主要表现在:动静脉血液完全分开、完全的双循环,心脏容量大,心跳频率快、动脉压高、血液循环迅速。
三、循环系统分类1.开管式循环:大多数无脊椎动物的血液循环系统都是“开放式”的,例如蝗虫的循环系统、虾的循环系统。
2.闭管式循环系统:所有的脊椎动物和部分无脊椎动物的循环系统是“封闭式”的,如蚯蚓、人类的循环系统。
3.二者区别a.开管式循环:是指动物体内的血液不完全在心脏与血管内流动,而能流进细胞间隙的循环方式.如节肢动物体内,背有心脏和它发出的血管(动脉)。
心脏两侧有具活瓣的心门,动脉直接开口在体腔。
心脏收缩时,心门关闭,血液从动脉的开口进入体腔,浸润各组织和器官。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
脊椎动物血液循环系统的演化
一:心脏
⒈位置:
心脏位于体腔前部,消化管腹侧的一个围心腔中,由围心膜所包被。
鱼类和有尾两栖类的围心腔位于体腔前方。
陆生脊椎动物的心脏向后,向腹方移动至体腔的前腹位。
⒉结构:
鱼类的心脏由静脉窦,一心房,一心室,动脉圆锥组成。
两栖类演变为两心房一心室,心房内出现完全或不完全的房间隔,静脉窦和动脉圆锥仍存在。
爬行类的心脏包括完全分隔的2个心房1个心室和退化的静脉窦,动脉圆锥消失,心室出现不完全分隔。
鸟类和哺乳类的心脏完全的分为四室,即左右心房和左右心室,其中哺乳类的左右心室之间有二尖瓣,左右心房之间有三尖瓣。
二:血液循环
鱼类的血液循环为单循环,即由心室压出的缺氧血经入鳃动脉进入鳃部进行气体交换,出鳃的多氧血不再回心脏而是经出鳃动脉直接沿背大动脉流到全身,从各组织器返回的缺氧学经主静脉系统再流回心脏,形成一个大圈。
两栖类为不完全双循环,左心房接受从肺静脉返回的多氧血,右心房接受从体静脉返回的缺氧血以及皮静脉返回的多氧血,它们最后均进入心室。
爬行类仍为不完全双循环。
鸟类和哺乳类为完全双循环,从体静脉回心的缺氧血经右心房进入右心室,被压入肺动脉弓,从肺静脉回心的多氧血经左心房进入左心室,被压入体动脉弓。
三:动脉系统
动脉系统的基本模式:腹大动脉,背大动脉,动脉弓胚胎期一般为六对
动脉弓的演变:
鱼类:由于以鳃呼吸,动脉弓在鳃部断裂为两部分,即入鳃动脉和出鳃动脉,中间以毛细血管联系,以进行气体交换。
软骨鱼类保留第2至6对动脉弓,硬骨鱼类保留第3至6 对动脉弓,其余退化。
两栖类以上的脊椎动物:成体因营肺呼吸,动脉弓不再断开,并只保留第3,4,6对动脉弓。
第三对为颈动脉,分布于头部和脑;第四对为体动脉弓,左右体动脉汇合成背大动脉;第六队为肺动脉弓;其中鸟类成体仅保留右体动脉弓,哺乳类则保留左体动脉弓。
四:静脉系统
⒈
鱼类:具H型主静脉系统,一对前主静脉窦,一对后主静脉窦,一对总主静脉,最后汇入静脉窦。
两栖类:四足类的基本模式—Y型大静脉(腔静脉)和肺静脉,出现一对前大静脉,一对后大静脉。
肺静脉的出现与肺的出现相适应,肺静脉直接进入左心房。
爬行类:肾门静脉趋于退化
鸟类:肾门静脉更趋退化,对提高后肢血液回心脏的血流速度和血压有积极意义。
哺乳类:进一步简化,肾门静脉完全退化消失,多数哺乳类仅保留右前大静脉。
⒉门静脉系统
门静脉两端为毛细血管,官腔内无瓣膜。
肝门静脉,肾门静脉和垂体门静脉系统。
脊椎动物静脉系统的演变趋势为:
a Y型大静脉系统代替了H型主静脉系统,静脉主干逐渐简化和集中。
b陆生脊椎动物出现了肺静脉,与肺脏的出现相呼应
c肝门静脉在各纲动物中均很稳定,保证营养代谢的需要
d肾门静脉由发达逐渐退化消失,提高回心血流的速度和血压。