重组和转座(32)
合集下载
分子生物学:重组和转座

转座子的特点
转座子是不必借助同源序列就可以移动的片断,即转座作用与供体 和受体的序列无关;
原核生物和真核生物都有转座子; 转座序列可沿染色体移动,甚至在不同染色体间跳跃(跳跃基因) 转座子对基因组而言是一个不稳定因素,它可导致宿主序列删除、
倒位或易位,并且其在基因组中成为“可移动的同源区”。位于 不同位点的两个拷贝转座子之间可以发生交互重组,从而造成基 因组不同形式的重排。 有些转座子与基因组的关系犹如寄生,它们的功能只是为了自身 的扩增与繁衍,因此被称为是自私的DNA。
转座中涉及的机制依赖于DNA 链的切割和重接,因此 (branch migration)
Cro蛋白抑制C I基因的转录,它占优势噬菌体即进入繁殖周期,并导致宿主细胞裂解。
与重组过程联系起来。 二者含有共同的核心序列15bp(O区)。
DNA转座子上携带作为重组位点的DNA序列,以及参与重组的蛋白质的基因(转座酶基因)。 同源重组 (homologous recombination )或普遍性重组(generalized recombination )
复制式转座 如 Tn3
非复制式转座 如 Tn10
类病毒反转录转座子/反转录病毒
带有反向终端重复序列.反向终端 重复序列嵌入在较长的正向排列的 重复序列(长末端重复序列, long terminal repeats LTR)中;
带有靶位点重复序列; 类病毒反转录转座子编码两种移位所需的蛋白:转座酶和反转录酶。 类病毒反转录转座子和反转录病毒的区别在于:反转录病毒的基因
径的歧化做好准备。
位点特异性重组与同源重组的区别
带有2个基因ORF1和ORF2;
同源重组 (homologous recombination )或普遍性重组(generalized recombination )
第五章 重组和转座

• 整合反应由λ噬菌体int基因的产物整合酶
(integrase, Int)催化。Int是一种DNA结 合蛋白,对POP ’ 序列有强的亲和力。 • 整合反应还需要一种有大肠杆菌编码的一 种细菌蛋白,称为整合宿主因子IHF (integration host factor, IHF) 。 • Int和IHF可以在体外进行位点特异性重组。
单链侵入模型:异源双链首先只在两个DNA分子中的一个形成。 Holliday中间体形成以后通过支链迁移能够在另一个DNA分子上产 生异源双链,这样就解释了异源双链是如何形成的。
双链断裂模型
二、异源双链和基因转换:
• 同源性重组时,在两个DNA分子之间互补碱
基配对的区域称为异源双链区 (heteroduplex region)。 • 由于异源双链区存在不配对碱基,两条链 就会在不配对部位发生错配。修复与否, 可以通过子囊菌减数分裂时孢子的分离情 况加以判断。
•
Insertion sequences have inverted terminal repeats and generate direct repeats of flanking DNA at the target site. In this example, the target is a 5 bp sequence. The ends of the IS consist of inverted repeats of 9 bp, where the numbers 1 through 9 indicate a sequence of base pairs.
3.Tn A家族:
• TnA是复制型转座的转座子,长约5kb左右 。 • 两端具有末端反向重复序列(而不是IS),长约38bp左右, •
重组和转座32ppt课件

Replicative transposition involves two types of enzymatic activity:
转座酶:transposase 解离酶:Resolvase
2. 非复制型转座 (nonreplicative transposition)
Nonreplicative transposition allows a transposon to move as a physical entity from a donor to a recipient site. This leaves a break at the donor site, which is lethal unless it can be repaired.
“We are sadly ignorant of the organization of the chromosome and of the possible types of changes in this that may occur to the chromosome as a whole or at the locus level”.
四、模板选择(copy choice)性重组
适用于RNA病毒,在这种重组中,聚合酶 从一个模板转换到另一个模板来合成RNA, 结果新合成的分子将含有两个不同亲本的遗 传信息
第二节 同源重组
第三节 转座(transposition)
一、原核转座子的类型
1.插入序列(insertion sequences , IS): IS家族的结构:
靶DNA,再将转座子连接到靶DNA的凸出单链上,最后填 补空缺完成转座。
The direct repeats of target DNA flanking a transposon are generated by the introduction of staggered cuts whose protruding ends are linked to the transposon.
转座酶:transposase 解离酶:Resolvase
2. 非复制型转座 (nonreplicative transposition)
Nonreplicative transposition allows a transposon to move as a physical entity from a donor to a recipient site. This leaves a break at the donor site, which is lethal unless it can be repaired.
“We are sadly ignorant of the organization of the chromosome and of the possible types of changes in this that may occur to the chromosome as a whole or at the locus level”.
四、模板选择(copy choice)性重组
适用于RNA病毒,在这种重组中,聚合酶 从一个模板转换到另一个模板来合成RNA, 结果新合成的分子将含有两个不同亲本的遗 传信息
第二节 同源重组
第三节 转座(transposition)
一、原核转座子的类型
1.插入序列(insertion sequences , IS): IS家族的结构:
靶DNA,再将转座子连接到靶DNA的凸出单链上,最后填 补空缺完成转座。
The direct repeats of target DNA flanking a transposon are generated by the introduction of staggered cuts whose protruding ends are linked to the transposon.
第二十三章重组和转座
第七节 真核生物的转座因子
一、玉米中的控制因子
1938年Marcus Rhoades首次发现不稳定突变等
位基因(unstable mutant allele),即一种回 复突变率很高的等位基因。不稳定是取 决于不连锁的Dt基因的存在。
McClintock。1940~1950描述了大量的控制因子
A1 A1 dt dt
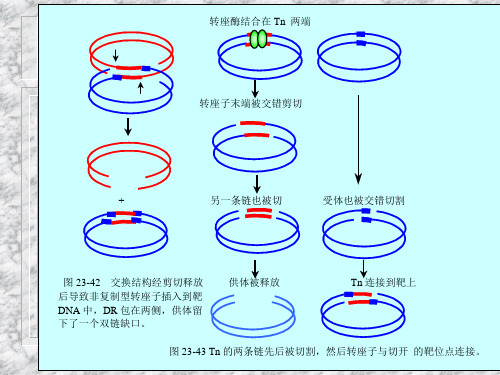
转座酶结合在 Tn 两端 转座子末端被交错剪切
+
另一条链也被切
受体也被交错切割
图 23-42 交换结构经剪切释放 后导致非复制型转座子插入到靶 DNA 中,DR 包在两侧,供体留 下了一个双链缺口。
供体被释放
Tn 连接到靶上
图 23-43 Tn 的两条链先后被切割,然后转座子与切开 的靶位点连接。
复制型转座模型解释了 (1) 复制性转座在转座后原来的位置上保留原
LTR 反转录病毒 gag
LTR pol int env
δ gag int
pol
δ
Ty
TyA
TyB
LTR gag int
pol LTR
Copia
ORF
pol
L1
ORF1
ORF2
图 23-72 病毒家族的反转录子末端有重复顺序内部有 开放读框
无LTR的反转录转 座子通过切开靶位 点双链,提供了引 物末端。反转录转 座子作为模板合成
Copia(果蝇)
Pol Ⅲ 转录 的加 工假 基
因
LINES L1(哺乳动物)
末端
长末端重复序列
无末端重复序列
靶重复序列
4~6 bp
7~12bp
阅读框
反转录酶和/或整合酶 无(不编码和转座有关
重组和转座

细菌的attl位点称为att B,其序 列的成份是BOB′,在噬菌体上的att 位点,称为att P,含有POP′。“O” 序列是att B和att P共有的,被称为 核心(core)序列,长15bp,重组 就发生在此序列上。两侧序列B,B′ 和P,P′是作为臂(arms);臂序列 各不相同。 在att位点的整合和切离并不涉 及反应序列的同源部分。整合需识 别att P和att B;而切离时需要att L和 att R。 整合和切离这两个反应所依赖 的蛋白不完全相同。整合(att B ×att P)反应需要噬菌体int基因的 产物和宿主的整合因子(integration host factor,IHF)。切离(att L × att R)反应需要噬菌体Xis基因的产 物以及Int和IHF蛋白的参与。两个 反应都需要Int和IHF,而Xis在控制 方向上起到重要的作用。它是切离 所需要的,对整合起抑制作用。
四、模板选择(copy chioce)性重组,适用 于RNA病毒,在这种重组中,聚合酶从一 个模板转换到另一个模板来合成RNA,结 果新合成的分子将含有两个不同亲本的遗 传信息
第二节 同源重组
一、Holliday连接: 1. 在同源染色体的相 同位点产生切口 2. 两条染色体的DNA 链发生交叉 3. 切口封闭,形成 holliday junction(或chi结构) 4. Branch migration (分枝迁移) 5. 在两对同源链的其 中任意一对上产生 切口,拆分中间体
非复制转座的断裂和 再连接过程: 在非复制转座方式中, “ strand transfer complex ”中 供体的未 断裂链被切开,转座 子两侧的靶链则被连 接
五、TnA家族转座需要转座酶和解离酶
第二十三章重组和转座

第二十三章重组和转座
•A1: 控制色素形成 •Dt: 控制产生斑点
第二十三章重组和转座
Marcus Rhoades分析了一种墨西哥玉米的穗, 此穗来自于一种籽粒有颜色的纯种自花受粉的玉米, 但它在后代中表现出一种意外的双因子杂种修饰性 孟德尔分离比 的植原物来,的自品交系产可生能了为上A1A述1d比td例t,。突变后产生A1a1Dtdt
第二十三章重组和转座
2020/12/9
第二十三章重组和转座
u 复制型转座模型解释了 u (1) 复制性转座在转座后原来的位置上保留原
有的Tn; u (2) 在新位置上转座子的两端出现正向重复靶
序列; u (3)转座过程中出现共合体。
第二十三章重组和转座
六.非复制型转座 七.Tn10的转座具有多项控制 u 1.“ 多拷贝抑制”(multicopy inhibition)
第二十三章重组和转座
u 这样a1成为首次发现的不稳定突变等位 基因(unstable mutant allele)的例子。即一 种回复突变率很高的等位基因。然而这种 等位基因的不稳定是取决于不连锁的Dt基 因的存在。一旦回复突变的发生,它们就 变得稳定了;即Dt基因能离开A1基因,这时 A1表型不再改变。这样a1表型是由一个缺陷 型的转座因子的插入而产生也就顺理成章 了(缺陷的转座因子自己并不能移动)。 Dt的缺乏使得表型保持稳定。
•
第二十三章重组和转座
第二十三章重组和转座
第二十三章重组和转座
第八节 反转录病毒和反转录子
u 一.反转录病毒(retroviruses) u (一) 反转录病毒的生活史 u 反转录子(retroposons) u 反转录转座子(retrotransposons) u 1. 反转录病毒基因组的结构与功能 u 2. 反转录病毒的整合模型 u 3. 反转录病毒可转录细胞的序列 u 二.酵母的Ty因子(transponson yeast)
•A1: 控制色素形成 •Dt: 控制产生斑点
第二十三章重组和转座
Marcus Rhoades分析了一种墨西哥玉米的穗, 此穗来自于一种籽粒有颜色的纯种自花受粉的玉米, 但它在后代中表现出一种意外的双因子杂种修饰性 孟德尔分离比 的植原物来,的自品交系产可生能了为上A1A述1d比td例t,。突变后产生A1a1Dtdt
第二十三章重组和转座
2020/12/9
第二十三章重组和转座
u 复制型转座模型解释了 u (1) 复制性转座在转座后原来的位置上保留原
有的Tn; u (2) 在新位置上转座子的两端出现正向重复靶
序列; u (3)转座过程中出现共合体。
第二十三章重组和转座
六.非复制型转座 七.Tn10的转座具有多项控制 u 1.“ 多拷贝抑制”(multicopy inhibition)
第二十三章重组和转座
u 这样a1成为首次发现的不稳定突变等位 基因(unstable mutant allele)的例子。即一 种回复突变率很高的等位基因。然而这种 等位基因的不稳定是取决于不连锁的Dt基 因的存在。一旦回复突变的发生,它们就 变得稳定了;即Dt基因能离开A1基因,这时 A1表型不再改变。这样a1表型是由一个缺陷 型的转座因子的插入而产生也就顺理成章 了(缺陷的转座因子自己并不能移动)。 Dt的缺乏使得表型保持稳定。
•
第二十三章重组和转座
第二十三章重组和转座
第二十三章重组和转座
第八节 反转录病毒和反转录子
u 一.反转录病毒(retroviruses) u (一) 反转录病毒的生活史 u 反转录子(retroposons) u 反转录转座子(retrotransposons) u 1. 反转录病毒基因组的结构与功能 u 2. 反转录病毒的整合模型 u 3. 反转录病毒可转录细胞的序列 u 二.酵母的Ty因子(transponson yeast)
重组的类型机制与转座

对进化生物学的影响
揭示物种进化的机
制
重组和转座是物种进化的重要驱 动力之一,研究它们有助于深入 了解物种进化的机制和过程。
解析生物多样性的
形成
重组和转座在生物多样性形成中 发挥重要作用,通过研究它们可 以更好地解析生物多样性的形成 机制和过程。
探索生命起源和演
化
通过研究重组和转座,可以更深 入地探索生命起源和演化的奥秘 ,了解生命的本质和演化规律。
。
转座酶
转座子移动需要转座酶的作用 ,这些酶能够切割转座子与 DNA的连接点,并将其插入
新的位置。
转座机制
转座子可以插入到基因内部或 基因间,影响基因的表达和功 能,也可能导致基因突变和重
排。
03
转座
转座的分类
转座子
可自主复制的转座元件,能在基因组内不同位置复制 和移动。
逆转座子
需依赖病毒或质粒进行复制的转座元件,通常从RNA 转录本中转录出cDNA并插入基因组。
THANKS
感谢观看
对医学和生物技术的意义
疾病发生机制的研究
重组和转座与许多疾病的发生和发展密切相关,研究它们有助于深 入了解疾病的发生机制和发展过程。
药物设计和治疗策略的制定
通过研究重组和转座,可以发现新的药物靶点和治疗策略,为药物 设计和治疗提供新的思路和方法。
生物技术的开发和利用
重组和转座是生物技术研究和应用的重要领域之一,通过研究它们 可以促进生物技术的开发和利用,推动相关领域的发展。
重组的类型、机制与转座
目 录
• 重组的类型 • 重组的机制 • 转座 • 重组与转座的关系 • 重组与转座的研究意义
01
重组的类型
同源重组
DNA的重组与转座

6
根据不同的机制,可将重组分成4类: • 同源性重组(homologous recombination) • 位点特异性重组(site-specific
recombination) • 异常重组(illegitimate recombination) • 转座重组(transposition recombination)
• Holliday连接体也能通过碱基之间氢键的断裂 和再连接而发生左右移动。这个过程称为支链 迁移(branch migration)。
24
25
27
• Holliday认为在DNA分子上存在某些位点,特 殊的引发重组的酶能够识别这些位点,确保两 条链在相同的部位被切断。
• 目前还没有足够的证据证明这些位点的存在。
17
18
20
• 一旦Holliday连接体形成后,它能进行重排从 而改变链的彼此关系。这种重排称为异构化, 因为在此过程中没有键的割裂。
• 一旦形成Holliday连接体后,就能被拆分。是 否发生重组依赖于拆分时Holliday连接体的构 象。
19
20
22
21
23
22
24
23
25
• Holliday模型被称为双链侵入模型,因为由于 每一个DNA分子的一条链侵入到另一个DNA 分子,它解释了在重组时两个DNA分子的异源 双链是如何形成的。
10
• 真核生物减数分裂时的染色体之间的交换, 某些低等真核生物及细菌的转化、转导、 接合,噬菌体的整合等都属于同源性重组 这一类型。
• 在整个基因组中,同源重组的频率并不恒 定,并且跟染色体的结构有关。例如在异 染色体附近遗传物质的交换要受到抑制。
11
1. 进行同源重组的基本条件
根据不同的机制,可将重组分成4类: • 同源性重组(homologous recombination) • 位点特异性重组(site-specific
recombination) • 异常重组(illegitimate recombination) • 转座重组(transposition recombination)
• Holliday连接体也能通过碱基之间氢键的断裂 和再连接而发生左右移动。这个过程称为支链 迁移(branch migration)。
24
25
27
• Holliday认为在DNA分子上存在某些位点,特 殊的引发重组的酶能够识别这些位点,确保两 条链在相同的部位被切断。
• 目前还没有足够的证据证明这些位点的存在。
17
18
20
• 一旦Holliday连接体形成后,它能进行重排从 而改变链的彼此关系。这种重排称为异构化, 因为在此过程中没有键的割裂。
• 一旦形成Holliday连接体后,就能被拆分。是 否发生重组依赖于拆分时Holliday连接体的构 象。
19
20
22
21
23
22
24
23
25
• Holliday模型被称为双链侵入模型,因为由于 每一个DNA分子的一条链侵入到另一个DNA 分子,它解释了在重组时两个DNA分子的异源 双链是如何形成的。
10
• 真核生物减数分裂时的染色体之间的交换, 某些低等真核生物及细菌的转化、转导、 接合,噬菌体的整合等都属于同源性重组 这一类型。
• 在整个基因组中,同源重组的频率并不恒 定,并且跟染色体的结构有关。例如在异 染色体附近遗传物质的交换要受到抑制。
11
1. 进行同源重组的基本条件
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
The direct repeats of target DNA flanking a transposon are generated by the introduction of staggered cuts whose protruding ends are linked to the transposon.
转座酶:transposase 解离酶:Resolvase
2. 非复制型转座 (nonreplicative transposition) Nonreplicative transposition allows a transposon to move as a physical entity from a donor to a recipient site. This leaves a break at the donor site, which is lethal unless it can be repaired.
第三节 转座(transposition)
一、原核转座子的类型
1.插入序列(insertion sequences , IS): IS家族的结构:
短的正向重复序列(direct repeats, DR) 略长的反向重复序列(inverted repeats,IR) 1kb左右的编码区,仅编码和转座有关的转座酶。
三、转座:
refers to the movement of a transposon to a new site in the genome. A transposon (transposable element) is a DNA sequence able to
insert itself (or a copy of itself) at a new location in the genome, without having any sequence relationship with the target locus. 转座分为复制型、非复制及保守型三种类型
二、Rec BCD途径 -同源重组的酶学机制
①
② ③ ④
⑤ ⑥ ⑦
Rec BCD蛋白同时具有解螺 旋酶的活性,使chi位点附近 切口的DNA解链 单链区被Rec A蛋白和SSB蛋 白覆盖 RecA 蛋白使单链DNA取代 双链DNA中的同源部分 D-loop区域的DNA产生切口, 新切口的3’端( the tail of newly nicked DNA )与另一 条DNA的单链区互补配对 DNA连接酶封闭切口,形成 Holliday junction RuvA 和RuvB发动迁移反应 RuvC拆分重组中间体
Barbara McClintock
“…that was hardly possible before the science of molecular biology caught up with McClintock”
Joshua Lederberg (1925-2008 ,1958 nobel Laureate )
Hale Waihona Puke 2. 复制型转座:① 在转座子和靶位点两端 分别交错切割产生切口; ② 转座子和靶位点的切口 末端交互连接(a-f, g-d), 形成一种交换结构 (cross structure); ③ 以游离的3’末端作为引 物进行复制,产生一个 包含两个正向重复的转 座子拷贝的复合物,称 为共合体(cointegrate), 这一过程在转座酶作用 下进行;
若玉米带有野生型C基因,则胚乳呈紫色; C基因的突变阻断了紫色素的合成,那么胚乳呈
白色; 在胚乳发育的过程中,突变发生回复导致斑点的 产生,回复突变发生在早期发育阶段,紫斑就比较 大; McClintock推测原来的C突变(无色素)是由一 个“可移动的控制因子”引起的,称解离因子 (dissociator,Ds),它可以插入到C基因中(即 转座)。 另一个可移动的控制因子是激活因子(activator, Ac),它的存在可激活Ds转座,进入C基因或其 他基因,也能使Ds从基因中转出,使突变基因产 生“回复突变”,这就是Ac-Ds系统
广泛一致性; 同源性重组可以发生在两条同源DNA上 的任何位点; 经常发生于减数分裂期的同源染色体之 间。
二、位点特异性重组
Site-specific recombination (Specialized recombination) 这种重组的特点是重组发生在特异位点,此位点含有短的 同源序列,供位点特异性重组酶识别(如 λ 噬菌体DNA整合 到寄主DNA); 重组DNA的其它区域不具有同源性
重组和转座
第一节 重组的分类
一、 同源重组或普遍性重组:
Recombination involving reaction between
homologous sequences of DNA is called generalized or homologous recombination.
依赖于参与重组的DNA分子在序列上的
3.Tn A家族:
TnA是复制型转座的转座子,长约5kb左右 。
两端具有末端反向重复序列(而不是IS),长约38bp左右,
任一个缺失都会阻止转座。 中部的编码区编码三个基因:转座酶,解离酶和抗性基因, TnA家族都带有抗性标记 。 靶位点具有5bp的正向重复序列。 解离位点(res)是TnA家族特有的内部位点。
④ 转座子的两个拷贝在 res(site of resolution )位点发 生重组,释放两个复 制子。这一过程称为 拆分(Resolution), 在解离酶的作用下进 行。
第四节 真核生物的转座因子
Barbara McClintock (芭芭拉· 麦克林托克) 1902-1992
一、 Ac-Ds系统
四、模板选择(copy choice)性重组
适用于RNA病毒,在这种重组中,聚合酶从 一个模板转换到另一个模板来合成RNA,结 果新合成的分子将含有两个不同亲本的遗传 信息
第二节 同源重组
一、Holliday连接: 1. 在同源染色体的相同 位点产生切口 2. 两条染色体的DNA 链发生交叉 3. 切口封闭,形成 holliday junction (或chi结构) 4. Branch migration (分枝迁移) 5. 在两对同源链的其中 任意一对上产生切口, 拆分中间体
二.转座机制:
根据转座子的机制,转座可分成三种不同的类型
1. 复制型转座 (replicative transposition):
Replicative transposition describes the movement of a
transposon by a mechanism in which first it is replicated, and then one copy is transferred to a new site. Replicative transposition involves two types of enzymatic activity:
对靶位点的选择有三种形式:随机选择,热点选择
和特异位点的选择。
Insertion sequences have inverted terminal repeats and generate direct repeats of flanking DNA at the target site. In this example, the target is a 5 bp sequence. The ends of the IS consist of inverted repeats of 9 bp, where the numbers 1 through 9 indicate a sequence of base pairs.
“We are sadly ignorant of the organization of the chromosome and of the possible types of changes in this that may occur to the chromosome as a whole or at the locus level”.
3.保守转座(conservative transposition) Conservative transposition involves direct movement with no loss of nucleotide bonds
三、转座中的一般过程:
1.非复制转座: 转座子插入到DNA上新的位点,首先交错切开靶DNA, 再将转座子连接到靶DNA的凸出单链上,最后填补空缺完 成转座。
2.复合转座子 (Composite transposons ):
含有一个中心序列和位于两侧的臂(arm); 除了和自身转座有关的基因外,中心序列含有
抗药性基因等遗传信息; 复合转座子两端的臂由IS序列组成。
A composite transposon has a central region carrying markers (such as drug resistance) flanked by IS modules.