过氧化氢对拟南芥根生长的影响
4.5生物拔高

4.5生物拔高题一、选择题:1.细胞是生物体结构和功能的基本单位,下列关于细胞的各项说法中不.正确的是( )A.洋葱根尖细胞有丝分裂过程中,核DNA分子数/染色体数的比值不能保持恒定B.成熟的植物细胞吸水膨胀,导致细胞液的渗透压降低C.胰蛋白酶从胰腺细胞排出,既需要消耗能量又需要细胞膜上载体蛋白的参与D.干细胞是一类具有分裂、分化能力的细胞2.下图中甲、乙、丙分别表示真核细胞内三种物质的合成过程,下列选项中正确的是( )A.丙过程合成物质的单体在生物体内约20种,该过程需要2种RNA参与B.甲、乙、丙过程中形成的核苷酸对种类分别为2、3、2C.在造血干细胞中甲、乙、丙过程均可发生D. 一个细胞周期中,甲乙过程在每个起点均可起始多次3.下列有关生物遗传、变异与进化的说法,正确的是( )A.由普通西瓜培育三倍体西瓜的原理是染色体变异,三倍体西瓜无籽性状不可遗传B.科学家通过基因工程育种,培育出了生产青霉素的高产菌株C.若在瓜田长期使用某种杀虫剂,导致害虫瓜蚜抗药基因频率增加,该田最终出现瓜蚜新物种D. 基因突变和基因重组都属于可遗传变异,两者都为生物进化提供了原材料4.下列关于生物体生命活动的调节过程正确的是()A.体液调节是通过体液传送激素的方式对生命活动进行的调节;B.免疫系统通过它的防卫功能、监控功能来实现其在维持稳态中的作用C.兴奋是以神经冲动的形式在神经元之间传递D.植物体的各个部位都能合成乙烯,其主要作用是促进果实成熟5. 现有A、B两个密闭透明的生态瓶,其生物组成和光照条件见下表。
下列叙述正确的A.两个生态瓶中的微生物、浮游藻类、水草和浮游动物共同组成了一个生物群落B.A瓶应放置在通风且光线良好,但避免阳光直射的位置C.实验结果发现A瓶持续时间比B瓶长,原因是A瓶的物质和能量能自给自足D.两组生态瓶进行对比实验,说明光照是所有生态系统必需的成分6.生物学是一门以实验为基础的自然科学。
下列与教材实验有关的叙述,正确的是( ) A .用健那绿和吡罗红混合染色剂将口腔上皮细胞染色,可观察DNA 和RNA 在细胞中的分布B .可用洋葱鳞片叶为实验材料提取类胡萝卜素和叶绿素C .用显微镜观察洋葱根尖临时装片时,需保持细胞活性以便观察有丝分裂过程D .鉴定待测样液中的蛋白质时,先加0.1g/ml 的NaOH 溶液1ml,摇匀后再加0.01g/ml CuSO 4溶液4滴29.(10分)富硒紫番薯含有丰富的微量元素和维生素,尤其是硒的含量较高。
过氧化氢参与了脱落酸调控的拟南芥根形态发育
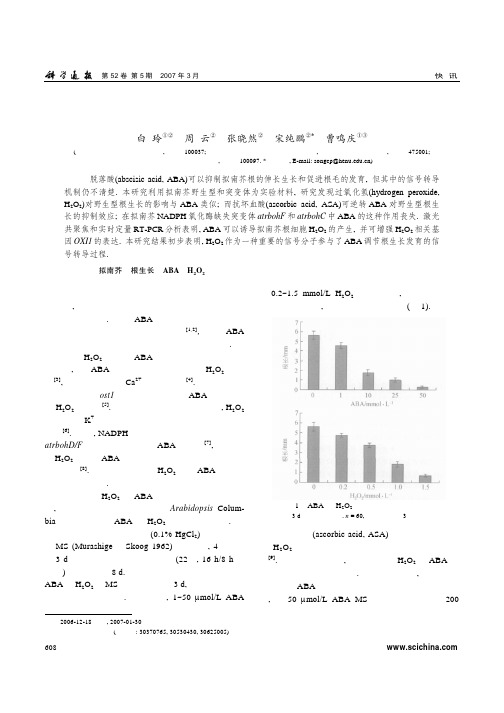
第52卷 第5期 2007年3月快 讯过氧化氢参与了脱落酸调控的拟南芥根形态发育白 玲①② 周 云② 张晓然② 宋纯鹏②* 曹鸣庆①③(① 首都师范大学生命科学学院, 北京 100037; ② 河南省植物逆境生物学重点实验室, 河南大学生命科学学院, 开封 475001;③ 北京农业生物技术研究中心, 北京 100097. * 联系人, E-mail: songcp@)摘要 脱落酸(abscisic acid, ABA)可以抑制拟南芥根的伸长生长和促进根毛的发育, 但其中的信号转导机制仍不清楚. 本研究利用拟南芥野生型和突变体为实验材料, 研究发现过氧化氢(hydrogen peroxide,H 2O 2)对野生型根生长的影响与ABA 类似; 而抗坏血酸(ascorbic acid, ASA)可逆转ABA 对野生型根生长的抑制效应; 在拟南芥NADPH 氧化酶缺失突变体atrbohF 和atrbohC 中ABA 的这种作用丧失. 激光共聚焦和实时定量RT-PCR 分析表明, ABA 可以诱导拟南芥根细胞H 2O 2的产生, 并可增强H 2O 2相关基因OXI1的表达. 本研究结果初步表明, H 2O 2作为一种重要的信号分子参与了ABA 调节根生长发育的信号转导过程.关键词拟南芥 根生长 ABA H 2O 22006-12-18收稿, 2007-01-30接受国家自然科学基金(批准号: 30370765, 30530430, 30625005)资助项目植物根系不仅是植物吸收水分和矿质营养的重要器官, 而且可以感知外界各种胁迫刺激从而调控植物的生长发育. 尽管ABA 作为一种重要的根源逆境信号对根生长发育的调控已有报道[1,2], 但是ABA 调节根生长发育信号转导的确切机制仍不甚了解.已知H 2O 2参与了ABA 调节植物生长发育的许多方面, 如ABA 可以诱导植物保卫细胞H 2O 2的产生[3], 并且调节质膜Ca 2+通道的活性[4]. 同时拟南芥气孔开放基因ost1编码的激酶介导了ABA 信号转导中H 2O 2的产生[5]. 在植物根发育的研究中发现, H 2O 2通过影响K +吸收相关基因的表达从而调控植物根的生长[6]. 同时, NADPH 氧化酶缺失的拟南芥双突变体atrbohD/F 幼苗根的生长不受ABA 的抑制[7], 从而推测H 2O 2可能是ABA 调控拟南芥根发育过程中的一个重要成分[8]. 但是目前还未见H 2O 2参与ABA 调控根发育的具体报道.为了研究H 2O 2在ABA 调控根生长发育中的功能, 我们首先利用野生型拟南芥Arabidopsis Colum- bia 为材料检测了ABA 和H 2O 2对根生长的影响. 野生型拟南芥种子经表面消毒(0.1% HgCl 2)后点种在无菌MS (Murashige 和Skoog 1962)培养基上, 4℃冷处理3 d 打破休眠后转入光照培养间(22℃, 16 h/8 h 光周期)垂直生长约8 d. 然后将幼苗转入含不同浓度的ABA 和H 2O 2的MS 培养基上生长3 d, 对根弯曲生长的长度进行测量统计. 研究表明, 1~50 µmol/L ABA与0.2~1.5 mmol/L H 2O 2的作用相似, 即都抑制拟南芥根的伸长生长, 且都具有浓度依赖性(图1).图1 ABA 和H 2O 2对拟南芥根生长的影响处理3 d 后根弯曲长度. n = 60, 标准误差由3次实验得到抗坏血酸(ascorbic acid, ASA)作为植物体内重要的H 2O 2清除剂确保了植物细胞内氧化还原态的平衡[9]. 借助于抗坏血酸, 我们研究了H 2O 2在ABA 调控拟南芥根生长中的作用. 研究分析表明, 抗坏血酸可以逆转ABA 对野生型拟南芥根伸长生长的抑制效应, 在50 µmol/L ABA MS 培养基上同时施加200快 讯第52卷 第5期 2007年3月mg/L 抗坏血酸后, 野生型根生长长度由0.3 mm 增至4.5 mm, 受ABA 抑制的程度明显减弱(图2); 同样1.0 mmol/L H 2O 2处理时拟南芥的根伸长平均为1.8 mm, 而和200 mg/L 抗坏血酸共同处理时增至3.4 mm(图2), H 2O 2抑制根 生长的现象也得到明显缓解, 进一图2 抗坏血酸逆转ABA 对拟南芥根生长的抑制效应(a) 分别在MS(CK), 50 µmol/L ABA, 1.0 mmol/L H 2O 2, 50 µmol/LABA + 200 mg/L 抗坏血酸, 1.0 mmol/L H 2O 2+200 mg/L 抗坏血酸培养基上培养3 d 的幼苗; (b) 处理3 d 后根弯曲长度. n = 60, 标准误差由3次实验得到步暗示了H 2O 2参与了ABA 调控的幼苗根生长.ABA 处理保卫细胞后可以诱导产生H 2O 2从而使气孔关闭[3,4]. 与此相似, 为了获得H 2O 2参与ABA 调控抑南芥根生长发育的直接证据, 我们以2,7-di- chlorofluorescin diacetate (H2DCF-DA)作为胞内H 2O 2的荧光探针[10,11], 采用激光扫描共聚焦显微成像(laser scan confocal microscope, LSCM)技术检测根细胞内H 2O 2含量的变化. 游离出的拟南芥根原生质体[12]在加入50 µmol/L H2DCF-DA (MERCK, San Diego, CA) 孵育20~30 min 后, 在LSCM (Bio-Rad MicroRadiance)下观察, LSCM 的工作条件为: E x = 488 nm, E m = 522 nm, 激光功率3%, 连续扫描记录时间360 s. 与对照(不加ABA 处理)相比, 50 µmol/L ABA 处理后拟南芥根细胞原生质体的DCF 荧光明显增强(图3(a)), 细胞中的平均荧光强度统计结果(图3(b))也明显表明ABA 可以诱导野生型拟南芥根细胞内H 2O 2的产生.我们又利用NADPH氧化酶缺失突变体atrbohF 和atrbohC 来对拟南芥根的生长发育进行研究. atr-bohF 和atrbohC 对ABA 不敏感, 在50 µmol/L ABA 的培养基上生长3 d 后根弯曲生长分别是相应野生型图3 ABA 促进拟南芥根细胞原生质体H 2O 2的产生(a) 50 µmol/L ABA 处理H2DCF-DA 孵育的根原生质体细胞LSCM 扫描荧光图, 标尺示10 µm; (b) 荧光强度平均数据统计图, n = 6第52卷 第5期 2007年3月快 讯的7和9倍多(图4). 这一结果进一步说明, H 2O 2的产生对于ABA 调节的拟南芥根早期生长发育是必须的.图4 H 2O 2 参与ABA 调控的拟南芥根的生长(a) 50 µmol/L ABA MS 培养基上生长3 d 的WT, atrbohC 和atrbohF ; (b) CK 及50 µmol/L ABA MS 培养基上培养3 d 的atrbohC 和atrbohF相对于WT 的相对根长. n = 60, 标准误差由3次实验得到为了进一步研究H 2O 2在ABA 调节的根发育中的功能, 我们对不同处理的根尖形态也做了分析. 结果表明, 50 µmol/L ABA 和1 mmol/L H 2O 2都可以诱导野生型拟南芥根毛的形成(图5(a)). 生长在50 µmol/L ABA 条件下的atrbohC 和atrbohF 与野生型截然不同, 野生型主根根尖丛生大量根毛而突变体根尖几乎没有根毛(图5(a)). 但是当施加外源1 mmol/LH 2O 2后突变体的根毛明显增多(图5(a)), 这表明H 2O 2参与了ABA 促进根尖根毛形成的过程, 参与了ABA 调节的根形态的发育. 我们的结果还发现当50 µmol/L ABA 和200 mg/L 抗坏血酸共同处理时, 野生型根尖根毛显著少于ABA 处理, 与突变体用ABA 处理得到的结果类似(图5(b)); 而同时抗坏血酸和H 2O 2处理的根毛数相对于H 2O 2处理也明显减少. 根尖形态研究结果进一步证实了H 2O 2在ABA 调节的拟南芥根发育的信号转导途径中的作用.拟南芥氧化胁迫诱导基因1(AtOXI1)编码的蛋白激酶OXI1属于AGC 家族蛋白, 在调控根毛发育中起到重要的作用. 该基因缺失的纯合突变体与野生型相比, 根毛数量和长度都明显减少; 并且OXI1在氧化猝发信号转导途径中具有重要的作用, 其基因的表达在体内直接受H 2O 2的诱导[13,14]. 采用实时定量RT-PCR 技术我们对幼苗根部AtOXI1的转录进行了分析, 100 µmol/L ABA 或ddH 2O(CK)处理生长8 d 的野生型拟南芥幼苗5 h 后, 提取根部总RNA, 反转录后在Rotor-Gene 3000(Corbett Research, Australia)上作实时定量PCR 分析(以Actin2作内参, 对样品中目标基因进行相对定量分析. 扩增AtOXI1的专一性引物为: 5′-GATTTGAAGCCAGACAATGTGAT-3′和 5′′-ACCAGTTAGATTCGGCGGTTTAGA-3′; Actin2专一引物为: 5′-TTCCTCATGCCATCCTCCGTCTT- 3′和5′-CAGCGATACCTGAGAACATAGTGG-3′, 实验重复3次), 结果表明, ABA 处理后AtOXI1的相对转录量升高(图6), 暗示H 2O 2相关的信号转导途径参与了ABA 对根生长的调节. 同时OXI1作为调节根毛发育的基因, 其相对高的表达与ABA 诱导大量根毛的形成也是一致的.图5 H 2O 2参与ABA 调节的拟南芥根毛生长(a) CK, 50 µmol/L ABA 和 1.0 mmol/L H 2O 2培养基生长3 d 的WT, atrbohC 和atrbohF 幼苗根尖形态, 标尺示50 µm; (b) WT 在200 mg/L 抗坏血酸, 50 µmol/L ABA+200 mg/L 抗坏血酸及1.0 mmol/L H 2O 2+200 mg/L 抗坏血酸培养基上生长3 d 的根尖形态, 标尺示50 µm快讯第52卷 第5期 2007年3月图 6 ABA 上调AtOXI1的转录n = 3综上所述, 我们认为H 2O 2作为一种重要的信号分子参与了ABA 对拟南芥根生长发育的调控, ABA 通过诱导根细胞内产生更多的H 2O 2抑制根的伸长生长, 并调控OXI1的表达促进根毛的形成和发育. 我们的实验结果对于揭示和理解ABA 调控根发育的机制具有重要的理论价值, 但是中间的详细细节还有待于进一步的研究.致谢 感谢张骁教授在成文过程中提出了宝贵的修改意见和建议.参 考 文 献1 Smet I D, Zhang H M, Inze D, et al. A novel role for abscisic acidemerges from underground. Trends Plant Sci, 2006, 11: 434—439 2 Chen C W, Yang Y W, Lur H S, et al. A novel function of abscisicacid in the regulation of rice (Oryza sativa L) root growth and de-velopment. Plant Cell Physiol, 2006, 47: 1—133 Zhang X, Zhang L, Dong F C, et al. Hydrogen peroxide is involvedin abscisic acid–induced stomatal closure in Vicia faba . Plant Physiol, 2001, 126: 1438—14484 Pei Z M, Murata Y, Benning G, et al. Calcium channels activatedby hydrogen peroxide mediate abscisic acid signalling in guard cells. Nature, 2000, 406: 731—7345 Mustilli A C, Merlot S, Giraudat J, et al. Arabidopsis OST1 proteinkinase mediates the regulation of stomatal aperture by abscisic acid and acts upstream of reactive oxygen species production. Plant Cell, 2002, 14: 3089—30996 Shin R, Schachtman D P. Hydrogen peroxide mediates plant rootcell response to nutrient deprivation. Proc Natl Acad Sci USA, 2004, 101: 8827—88327 Kwak J M, Mori I C, Pei Z M, et al. NADPH oxidase AtrbohD andAtrbohF genes function in ROS-dependent ABA signaling in Arabidopsis . J EMBO, 2003, 22: 2623—26338 Gapper C, Dolan L. Control of plant development by reactiveoxygen species. Plant Physiol, 2006, 141: 341—3459 Noctor G, Foyer C H. Ascorbate and glutathione: Keeping activeoxygen under control. Annu Rev Plant Physiol Plant Mol Biol, 1998, 49: 249—27910 Schopfer P. Hydroxyl radical-induced cell-wall loosening in vitroand in vivo : Implications for the control of elongation growth. Plant J, 2001, 28: 679—68811 Simontacchi M, Caro A, Fraga C G, et al. Oxidative stress affectstocopherol content in soybean embryonic axes upon imbibition and germination. Plant Physiol, 1993, 103: 949—95312 Demidchik V, Tester M. Sodium fluxes through nonselective cationchannels in the plasma membrane of protoplasts from Arabidopsis roots. Plant Physiol, 2002, 128: 379—38713 Anthony R G, Henriques R, Helfer A, et al. A protein kinase targetof a PDK1 signalling pathway is involved in root hair growth in Arabidopsis . J EMBO, 2004, 23: 572—58114 Rentel M C, Lecourieux D, Ouaked F, et al. OXI1 kinase is neces-sary for oxidative burst-mediated signaling in Arabidopsis . Nature, 2004, 427: 858—861。
四种药品处理对拟南芥dna甲基化的影响

obviously promoted plant root length of Arabidopsis seedlings. The analysis by MSAP showed that levels of 5-mdC in mannitol-treated seedlings were also altered significantly, ranging from 17.75%, 21.15%, 15.49% to 46.10% in plants treated with 50, 100, 150 and 200 mmol·L-1 mannitol, respectively. The results indicted that mannitol causes epigenetic variations in all the samples based on different levels and patterns of DNA methylation. With 50, 100, 150 and 200 mmol·L-1 mannitol treatment, methylation and demethylation of DNA were 5.78%, 15.48%, 10.71%, 33.73% and 10.98%, 5.36%, 8.33%, 7.69%. Mannitol causes a dose-dependent and transient change of global 5-methyldeoxycytidine levels in Arabidopsis seedlings.
(1) The effect of oxytetracycline stress on the development as well as genomic DNA methylation levels and patterns of Arabidopsis thaliana seedlings was assessed. Oxytetracycline at concentrations of 3, 5, 7, and 9 µmol·L-1 visibly inhibited root and stem enlongation of A. thaliana seedlings, but significantly promoted lateral root proliferation. Methylation sensitive amplified polymorphism (MSAP) analysis showed that genomic DNA methylation levels of oxytetracycline stressed seedlings were lower than that of the untreated plants. The results indicated that oxytetracycline stress induced epigenetic variations in A. thaliana seedlings; no clear trend or pattern of 5-methylcytosine percentage change emerged. At oxytetracycline concentrations of 3, 5, 7, and 9 µmol·L-1, methylation and demethylation of A. Thaliana seedling genomic DNA were respectively13.29%, 9.22%, 8.03%, 12.59% and 2.80%, 4.26%, 5.11%, 4.90%.
拟南芥生长发育和抗逆性的分子机理研究

拟南芥生长发育和抗逆性的分子机理研究植物生长发育及其抗逆性一直是植物学研究的热门领域。
拟南芥,因其生长周期短,遗传背景清晰,成为了模式植物,被广泛应用于该领域。
本文将着重介绍拟南芥生长发育及其抗逆性的分子机理研究进展。
一、拟南芥的基础生长发育拟南芥的生长发育可以分为两个主要阶段:萌芽期和成长期。
在萌芽期,拟南芥发芽后,主要为幼芽和根系发育,而在成长期,则是幼苗进一步生长、分化出花器官、形成花后,再逐渐形成果实。
1. 根系生长发育拟南芥根系的生长发育与环境条件密切相关。
根系统以分化出来的主根和分支根为主体,发育出不同的形态和特性,以适应环境。
主根在土壤中向下延伸,同时分支根则向周围扩散。
在根系结构发育完成后,拟南芥根系还能收集到水、养分等,以维持植物的生长发育。
2. 幼苗生长发育拟南芥的幼苗是指在萌芽期已经分化出根系的拟南芥,即幼芽和根系已经形成。
在幼芽萌发后,会不断分化出新的叶片,并形成具有不同的形态和功能。
此时,拟南芥的生长速度较快。
3. 花器官形成花器官是植物重要的生殖器官。
在花器官形成的过程中,拟南芥会逐渐形成雌雄蕊和花瓣等构成花朵。
拟南芥的花器官形成受到许多基因调控,这些基因调控可以影响植物的花器官鲜艳程度、形态、数量以及其功能。
4. 果实成熟果实的成熟和植物的生长发育密切相关。
拟南芥果实成熟的过程中,植物会不断积累营养物质,直到果实形成。
同时,植物器官的分化也会触发一系列信号传递机制,以调节植物对环境的响应能力。
二、拟南芥的抗逆性植物的生长发育面临着不同的环境压力,如环境温度、酸碱度、干旱、盐渍等。
而在生长发育过程中,拟南芥实现了对这些压力的适应。
这得益于其多种基因的调节,进而影响植物的抗逆性。
1. 盐胁迫盐胁迫是影响拟南芥生长发育的常见环境压力,它使得植物经受不住盐分的侵蚀。
为了适应盐胁迫环境,植物通过调节促进盐分排除的基因表达,以及在组织水分调节方面的功能作用,来增强其抗逆性。
同时,植物还会通过调节根系结构和激素分布等相关基因来适应新的环境条件。
2023届福建省厦门一中高三一模生物试题(解析版)

D、若根不水平放置,而是垂直放置,那么所有组不论根是否具有向重力性都会呈现垂直状态,就无法确定谁有向重力性谁没有,因此根不能垂直放置,应该水平放置,方便观察现象,D错误。
D.图戊中4个细胞的基因型最可能为AB、Ab、aB、ab
【答案】B
【解析】
【分析】图示分析:甲是MI中期,乙是MI后期,丙是MII中期,丁MII后期,戊是MII末期。
【详解】A、应取该植物的花药制成临时装片,才能观察到上面的图像,A错误;
B、图甲是MI中期、乙细胞是MI后期,故甲乙中含有同源染色体,其中甲细胞中有12对同源染色体,故有12个四分体,B正确;
【详解】A、检验项目是否正常应与该项目的参考范围对照,说明内环境稳态是相对的,通常会由于内外因素的影响使内环境的理化性质和化学成分处于相对稳定状态,A正确;
B、检查者血液中谷丙转氨酶和球蛋白偏高,说明检查者的肝细胞可能受损,也可能有病毒感染,但还需要进一步检测,B正确;
C、若抽血化验单所有检验项目都在参考范围内,则只能说明检测的项目处于相对稳定状态,其他未检测的项目并不清楚,因而只能说明该检查者的内环境可能维持了稳定状态,C错误;
故选C。
8.稻蝗主要以禾本科植物叶片为食,是水稻的重大害虫。为调查某农场内稻蝗跳蝻(稻蝗幼虫)的密度,研究人员每次调查后隔3天再调查一次,所得数据如表所示。下列相关叙述错误的是()
调查次数
第一次
第二次
第三次
H2O2介导的H2S产生参与干旱诱导的拟南芥气孔关闭

植物学报Chinese Bulletin of Botan ydoi:10.37241SP.J.1259.2012.00217·研究报告·H202介导的H2S产生参与干旱诱导的拟南芥气孔关闭王兰香,侯智慧,侯丽霞,赵方贵,刘新。
青岛农业大学生命科学学院,山东省高校植物生物技术重点实验室。
青岛266109摘要以野生型拟南芥(Arabidopsis tha/iana)及其突变体(a trbo hD、at rbohF、atr bohD IF、at/-cdes、atd-cd es)和过表达株系(O EL.C De s、O ED.CO e s)材料,利用药理学实验,结合分光光度法和激光共聚焦显微技术,探讨硫化氢(h yd ro g en sulfide,H2S)在干旱诱导的拟南芥气孔关闭中的作用及其与过氧化氢(hydrogen peroxide,H202)雕]关系。
结果表明,H2S清除剂次牛磺酸(hypotaurine,HT)及合成抑制剂氨氧基乙酸(aminooxy acetic acid,AOA)、羟胺(hydroxylamine。
NH20H)和丙酮酸钾(potasium pyruvate,C3H3K03)+氨水(ammonia,NH3)均可不同程度抑制干旱诱导的气孔关闭;干旱对OEL-CDes和0ED.cDes植株气孔关闭的诱导作用明显.而atl-cdes和atd-cdes叶片气孔对干旱胁迫反应的敏感性下降:干旱胁迫能明显增加拟南芥保卫细胞CH202水平及叶片dPH2S含量,提高D-/L-半胱氨酸脱巯基酶活性及基因表达量,而对突变体atrbohD、 atrbohF和atrbohD,F没有显著影响。
清除H202可减弱干旱胁迫对H2S含量和D-/L一半胱氨酸脱巯基酶活性的诱导效应。
研究结果表MR2S位于H202下游参与干旱诱导拟南芥气孔关闭的信号转导过程。
关键词拟南芥,干旱胁迫,过氧化氢。
2021年青岛二中高三生物下学期期中考试试卷及参考答案

2021年青岛二中高三生物下学期期中考试试卷及参考答案一、选择题:本题共15小题,每小题2分,共30分。
每小题只有一个选项符合题目要求。
1. 如图甲表示某生物膜结构,a、b、c表示物质跨膜运输的方式,图乙表示物质运输速率与细胞内外浓度差的关系。
则m、n表示的物质跨膜运输方式可分别对应图甲中的()A.b、aB.c、aC.a、bD.c、b2. 细胞核由核膜、染色质、核仁、核孔组成,下列有关叙述正确的是()A.核孔实现了细胞间的信息交流B.染色质主要由DNA和蛋白质组成C.核仁与DNA合成以及线粒体的形成有关D.核膜是单层膜,把核内物质与细胞质分开3. 将下列生物或细胞放入清水中,会吸水胀破的是()A. 洋葱表皮细胞B. 大肠杆菌C. 草履虫D. 人成熟红细胞4. 下列关于人体X、Y染色体的叙述,错误的是A.X、Y染色体可形成四分体B.X、Y染色体上的基因可在生殖细胞中表达C.X、Y染色体上不存在等位基因D.X、Y染色体上有与性别决定相关的基因5. 下图是探究某绿色植物光合速率的实验装置图,装置中的NaHCO3溶液可维持瓶内的CO2浓度相对稳定,将该装置放在20℃、一定光照条件下。
实验开始时,针筒的读数是0.2 mL。
毛细管内的有色液滴在A处。
30 min后,针筒的读数需要调至0.6 mL,才能使有色液滴维持在A处。
下列有关叙述错误的是()A.该实验过程中光合速率大于呼吸速率B.若测定植物的实际光合速率,还需要在暗处测定呼吸速率C.若将小烧杯中的NaHCO3溶液换成清水,则一段时间后光合作用会停止D.若用该装置探究该植物生长的最适温度,NaHCO3溶液浓度和光照强度等都是无关变量6. 下列不属于体液调节的特点是A.调节作用范围广泛B.通过体液运送调节物C.调节速度缓慢,作用时间长D.调节物都是由内分泌腺产生的7. 下图为绿色植物某细胞内发生的两个生理过程。
下列分析正确的是()A. 进行℃过程的细胞也能进行℃过程B. ℃过程产生的能量,大部分用于合成ATPC. ℃、℃过程中的还原性辅酶是同种物质D. ℃过程发生在线粒体内膜上,℃过程发生在叶绿体内膜上8. 下图所示为叶绿体中的某种结构及其上发生的物质和能量变化,有关叙述错误的是()A. 光反应在类囊体薄膜上进行B. 图中℃上有光合色素C. 图中光能转变成了化学能D.[H]、O2和ATP都能用于暗反应9. 细胞膜可以控制物质进出细胞,下列叙述不正确的是()A.细胞需要的营养物质可以从外界进入细胞B.细胞产生的代谢废物可以排出细胞C.抗体、激素等物质在细胞内合成后,分泌到细胞外D.环境中一切有害的物质都不能通过细胞膜进入细胞10. 某学生利用性状分离比的模拟实验探究人类红绿色盲的遗传规律,下列叙述中错误的是()A.需要准备三种不同颜色的彩球,分别标记B、b、Y表示X B、X b、YB.若甲桶内装有B、b两种彩球数量各10个,则乙桶内装有两种彩球数量均可超过10个,且必有一种是Y 彩球C.通过女性色盲与男性正常婚配这组模拟实验可得出人类红绿色盲具有隔代交叉遗传的特点D.每次抓取小球统计后,应将彩球放回原来的小桶内,重复抓取50—100次11. A TP是细胞内绝大多数生命活动的直接能源物质。
2023~2024年上海闵行区生物高考一模试卷

图12023学年第一学期高三年级学业质量调研生物学学科注意:1.本考试设试卷和答题纸。
作答必须涂或写在答题纸上,在试卷上作答一律不得分。
2.“单选”指每题只有一个选项为正确答案;“多选”指每题有两个或两个以上选项为正确答案;“编号选填”指每空有一个或多个选项为正确答案。
一、高原鼠兔(15分)青藏高原东部牧区属于高山草甸生态系统(图1), 主要植被是多年生草本植物。
广泛分布于此的植食性 小型哺乳动物高原鼠兔,主要利用植被高度较低、视 野开阔的草地生境。
野外收集该区域的香鼬和艾虎粪 便进行分析,发现高原鼠兔在这两种动物的食物中出 现频次非常高。
1.(1分)高原鼠兔取食、挖掘、排泄、刈割等活动会使草甸表面形成土丘和裸露斑块,经过一段时间恢复,裸斑上逐渐有植物生长,但植物盖度较低,称之为秃斑。
形成秃斑的过程属于______(初生/次生)演替。
2.(2分)以下关于高山草甸生态系统的描述,正确的是______。
(多选)A .信息双向传递B .能量双向流动C .受全球变暖影响D .存在能量倒金字塔研究人员采用堵洞盗洞法估测高原鼠兔的种群数量,计算公式为:种群数量=有效洞口数/洞口系数。
将待测区域内高原鼠兔的洞口全部填埋,连续3天每天记录被盗开的洞口数并计算平均值,即有效洞口数;在附近另一片较小的区域内测定有效洞口数后,用夹捕法捕尽区域内的高原鼠兔,计算有效洞口数与种群数量的比值,即洞口系数。
3.(2分)关于堵洞盗洞法的叙述,下列错误..的是______。
(单选) A .该方法依据的是数学中的抽样原理B .测有效洞口数时,连续3天均应于同一时间段记录C .每天记录好被盗开的洞口数后,均需要重新填埋D .幼年高原鼠兔尚不能掘土打洞,会导致估测数量偏大图24.(2分)如图2,较小区域B 内的有效洞口数为4个,共有高原鼠兔35只。
则待测区域A 内的高原鼠兔的种群数量大约是______只。
研究人员选取了处于同等放牧条件下的研究区域,以扰动最强处为中心向扰动弱的方向,根据有效洞口数将高原鼠兔扰动由强到弱划分为4组,探究了高原鼠兔不同强度扰动对高山草甸生态系统和放牧质量的影响,结果如下表1所示。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
过氧化氢对拟南芥根生长的影响
摘要过氧化氢作为一种超氧化物能对植物产生氧化胁迫。
把野生型拟南芥培养到根长约为1.5 cm,然后转移到浓度为0.2~2.0 mmoL/L的过氧化氢培养基上,隔24 h对根的长度进行测量,了解过氧化氢对拟南芥根生长的影响。
试验结果表明,当过氧化氢浓度为0.2 mmoL/L时,即对植物根生长造成抑制;当浓度为 2.0 mmoL/L时,则植物根生长完全受到抑制,表现为根生长停止。
关键词过氧化氢;拟南芥;根生长;影响
拟南芥(Arabidopsis thaliana)是十字花科拟南芥属植物,虽然没有经济价值,但具有生育期短、植株个体小及基因组小等特点,因而长期以来一直被用来作为分子生物学和传统遗传学研究的模式试验材料,作为高等植物中具有最少基因组的物种,在科学研究中的地位极为重要。
为了与国际接轨,近年来国内已引入了拟南芥作为试验材料。
室内培养不仅可以得到试验所需的材料,为拟南芥新品种的培养和种性的改良提供便利条件,掌握拟南芥市内培养技术对顺利开展植物发育生物学的研究具有重要意义。
全球由于环境胁迫给作物造成的品质下降,产量降低的损失是惊人的。
当作物生长的外在条件如温度、温度、土壤中的水分,盐浓度等发生急剧变化或当大气污染(如SO2、臭氧)、紫外线辐射、某些农药如Paraquat(一种光动除草剂)及病原体等作用于植物时,都会使植物体内产生大量的活性氧(Reactive Oxygen Species,ROS),形成氧化损伤。
这些比氧活泼的含氧化合物包括超氧根阴离子(O2-)、氢氧根离子(OH-)、羟自由基(OH)、过氧化氢(H2O2)等。
高浓度的H2O2会对细胞膜系统和细胞带来伤害和毒害[1-2],H2O2 是活性氧的一种,是细胞有氧代谢的产物。
通过对影响拟南芥根生长的过氧化氢浓度的研究,可以了解其对根生长的影响,从而促进农业生产的研究[3-7]。
1材料与方法
1.1试验材料
试验材料为拟南芥种子,30%过氧化氢溶液。
1.2试验方法
1.2.1种植根长约1.5 cm的拟南芥。
准备一些中号EP管,以每EP管80粒种子为单位,对拟南芥种子用氯气熏蒸法(4 mL的浓盐酸加入100 mL 5.2%的NaClO 溶液)灭菌12 h。
然后用0.15% agar溶液悬浮,种在含有1/2 MS培养基的玻璃培养皿上,每皿约45粒种子。
放入4 ℃冰箱中72 h。
从冰箱取出后垂直放置在20 ℃的组织培养室中,大约3 d后,根即可长至1.5 cm。
1.2.2用不同浓度过氧化氢进行筛选。
待1/2 MS培养基温度约为40 ℃时,加入对应的30%的过氧化氢溶液,使培养基浓度分别为0.2、0.4、0.6、0.8、1.0、1.2、1.4、1.6、1.8、2.0 mmoL/L。
然后把根生长至1.5 cm的拟南芥幼苗用镊子轻轻的移到这些培养基中。
每皿放拟南芥幼苗18根,上下2行排列,各8根。
移完幼苗后用封口膜把圆玻璃皿封上口,用黑色mark笔在每个幼苗的根尖处点点作标记。
每个浓度重复3次,即做3个相同的皿。
1.2.3记录根生长的长度。
以转移苗子后的时间为准,每24 h对根的转移后生长的新长度进行测量,精确到小数点1位。
同一皿的根长取平均值。
2结果与分析
从表1和图1可以看出,当过氧化氢浓度为0.2 mmoL/L时,其对植物根生长造成抑制;当浓度为0.8 mmoL/L时,(上接第19页)
抑制作用较为明显,根的长度约为正常值的1/2;而当浓度为2.0 mmoL/L时,植物根生长遭到完全抑制,表现为根生长停止。
3结论与讨论
试验结果表明,过氧化氢会对拟南芥生长造成较为明显的影响,当浓度较大时,会抑制拟南芥幼苗的根的生长。
掌握过氧化氢对拟南芥生长的影响的情况,可以进一步筛选抗氧化的拟南芥突变体,也可以对拟南芥进行种前恶劣条件的驯化,以提高抗氧化能力。
4参考文献
[1] SAIRAM R K,RAO K V,SRIV ASTA V A G C.Differential response ofwheat genotypes to long term salinity stress in relation to oxidative stress,antioxidant activity and osmolyte concentration[J].Plant Sci,2002(163):1037-1046.
[2] SAIRAM RK,TYAGI A.Physiology and molecular biology of salinity stress tolerance in plants[J].Curr Sci,2004(86):407-421.
[3] 张司南,高培尧,谢庆恩,等.镉诱导拟南芥根尖过氧化氢积累导致植物根生长抑制[J].中国生态农业学报,2010(1):136-140.
[4] 杨礼香,王正询,柯德森,等.拟南芥血红蛋白(CAtGLB1)与过氧化氢的相互作用[J].氨基酸和生物资源,2009,31(4):1-4,16.
[5] 何金环,李利红,宋纯鹏.活性氧不敏感型拟南芥的突变体对H2O2的响应[J].植物生理学通讯,2007,43(1):31-35.
[6] 于天昊,洪宇,谢庆恩,等.外源蔗糖、VC对铬诱导的拟南芥主根生长抑制和过氧化物积累的缓解作用[J].中国农学通报,2010,26(6):289-292.
[7] 白玲,周云,张晓然,等.过氧化氢参与了脱落酸调控的拟南芥根形态发育[J].科学通报,2007,52(5):608-611.。