神经生长因子受体TrkA在中枢神经系统的表达、分布和作用
脊髓损伤的药物治疗

脊髓损伤的药物治疗脊髓损伤的药物治疗徐州市中⼼医院拾坤【疾病概述】脊髓损伤(spinal cord injury,SCI)可发⽣在任何年龄,以20岁到49岁青壮年男性居多。
交通事故伤、坠落伤及暴⼒损伤是主要原因。
美国每年有10000新发病例,每年为此花费超过60亿美元。
据估计,全世界每年新发⽣脊髓损伤约50万⼈,⽽截瘫患者将达到250万⼈,给社会和个⼈造成了沉重负担。
脊髓损伤曾被称为“不可治愈的疾病”,随着现代科技的进步和医学的发展,脊髓损伤患者的死亡率从20世纪初期的50%左右降低到⽬前的6%,但脊髓损伤后神经功能的恢复程度仍不令⼈满意。
【病理⽣理】脊髓损伤由原发性损伤和继发性损伤组成,原发性损伤是由撞击、切割、出⾎压迫等机械外⼒导致的组织损伤,⼀般为不可逆损伤;继发性损伤是在脊髓原发性损伤的基础上由⽔肿、炎症反应、钙超载、缺⾎-再灌注等病理⽣理过程导致的组织的损害,这种损害在原发伤之后继续发展⼏天⾄⼏个⽉,多具可逆性及可调控性,对其控制⽔平直接影响到最终脊髓功能恢复程度。
【治疗】对于脊髓损伤患者,除及时有效地稳定脊柱、解除脊髓压迫等治疗以外,早期予以有效的药物治疗也不可忽视,脊髓损伤后的药物治疗⽬的主要是1、减少脊髓损伤后早期创伤、炎症等的有害作⽤;2、对脊髓再⽣的促进并阻⽌抑制因⼦的影响;3、⽀持轴突再⽣并促进其与靶器官的联系等。
当前基因治疗已成为研究的热点,但要真正应⽤于临床还需很长时间,所以对保护脊髓、减少脊髓继发性损害的⼀些现有的药物仍应引起⼤家重视。
【治疗药物】⼀、类固醇类1、甲基强地松龙(methylprednisolone,MP,甲强龙)甲强龙是⼀种合成的中效糖⽪质激素,其抗炎作⽤是氢化可的松的5倍,⾎浆半衰期为2.5⼩时,甲强龙由于临床⽤量不同⽽表现出不同的临床作⽤,其作为常规药物⽤于治疗脊髓损伤病⼈始于上世纪,是唯⼀被美国联邦⾷品药品管理局(Food Drug Administration,FDA)批准的脊髓损伤治疗药物。
神经生长因子

神经生长因子(NGF)成都医学院 2013级临床七班马成杰 1325100381摘要:神经生长因子(NGF)是神经营养因子中最早被发现,目前研究最为透彻的,具有神经元营养和促突起生长双重生物学功能的一种神经细胞生长调节因子,它对中枢及周围神经元的发育、分化、生长、再生和功能特性的表达均具有重要的调控作用。
NGF包含α、β、γ三个亚单位,活性区是β亚单位,由两个118个氨基酸组成的单链通过非共价键结合而成的二聚体,与人体NGF的结构具有高度的同源性,生物效应也无明显的种间特异性。
关键词:神经生长因子(NGF)分子组成生物效应1951年,Levi-M on talc in i和Hamburger首先在培养的小鼠肉瘤细胞内发现NGF存在。
从发现第一个多肽生长因子NGF以来,又相继发现了脑源性神经营养因子(brain-derived neurotrophic factor,BDNF)、神经营养素-3(neurotrophic-3,NT-3)、神经营养素-4(neurotrophic-4,NT-4)、神经营养素-5(neurotrophic-5,NT-5),由于它们起源于同一基因家族,故可称为NGF 家族。
此外,还有睫状神经营养因子(c i l i a r y neurotrophic factor,CNTF),胶质细胞源性神经营养因子(g l i a l-derived neurotrophic factor,GDNF),但它们在分子结构和受体类型上与NGF家族不同。
经典的NGF是从小鼠颌下腺中分离所得的分子量为140,000的糖蛋白,由α、β、γ三种肽链构成,肽链间以共价键结合。
其中β亚单位是NGF的活性区。
经过40多年的研究,NGF的蛋白结构已基本了解清楚,三维结构已从其晶体图像证实。
它由两个相同的亚单位组成,亚单位由三个β转角连接的三对反平行β折叠组成。
亚单位间通过“半胱氨酸结点”连接。
目前发现的NGF均为糖蛋白,其构型至关重要的氨基酸在神经营养素家族中保守,不同于NGF的特性由可变区的氨基酸决定,可变区的变化不影响它们的基本结构,但对特异性受体的结合起决定作用。
31中枢神经系统活动基本规律-精选文档
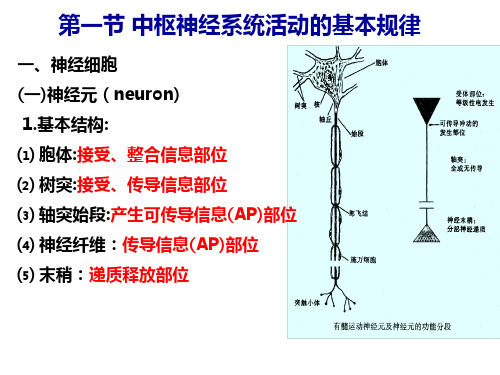
五、神经系统功能的基本方式 (一)反射与反射弧
1.反射(reflex):在CNS参与下,机体对内外环境刺激 的规律性应答反应。 2.分类:
条件反射 非条件反射 3.反射弧: 反射弧(reflex arc)是反射的结构基础和基本单位。
感受器→传入N→中枢→传出N→效应 器
4.反射过程:
通透性↑
Cl-(主) K+
通透性↑
EPSP
IPSP
4.化学突触传递的特征:
⑴单向传递:突触前神经元→突触后神经元。 ⑵突触延搁:需时0.3~0.5ms/个突触。 ⑶总和:时间总和和空间总和。 (4)对内环境变化的敏感性:对缺氧、PCO2↑、
药物敏感(如pH↑→N元兴奋性↑;士的宁→递质 释放↓;咖啡因→递质释放↑)。
习惯化:重复刺激时,突触对刺激的反应逐渐减 弱或消失。 敏感化:与习惯化相反。 长时程增强(LTP):短时间内快速重复刺激后,突 触后N元产生一种快速形成的和持续性的突触后电 位增强(持续时间大于强直后增强)。 长时程抑制(LTD):与LTP相反。
四、中枢神经递质和受体
(一)中枢神经递质(transmitter)
(5)易疲劳性:与递质的耗竭有关。
(二)电突触传递
结构基础:缝隙连接
二个N元紧密接触的部位上有水通道蛋 白,允许带电离子通过,且电阻低。
传递过程:电-电(以局部电流方式)。 传递特征:双向性,速度快,几乎
无潜伏期。
(三)非突触性化学传递
结构基础:轴突末梢分支 上有结节状的曲张体,曲 张体内含有递质小泡。
二、兴奋传递的方式
突触(synapse):一个神经 元与另一个神经元或其他细 胞相接触的部位。
类型:化学突触传递 电突触传递 非突触性化学传递
NGF_BDNF及受体trkA_trkB_trkC在正常猴脊髓的表达

Sep.2002 18(3) 240~242神经解剖学杂志(Chinese Journal of Neuroanatomy)NGF、BDNF及受体trkA、trkB、trkC在正常猴脊髓的表达¹伍校琼 李昌琪 刘 丹 蔡维君 罗学港(中南大学湘雅医学院 神经生物学研究室,长沙410078)摘 要 采用免疫组织化学方法观察了神经生长因子(NGF),脑源性神经营养因子(BDNF)以及NGF家族因子受体trkA、trkB、trkC的免疫阳性反应在正常猴脊髓的分布。
结果表明:NGF免疫反应阳性的神经元在脊髓灰质各层中均有分布,灰、白质内也可见较多的NGF免疫反应阳性的胶质细胞。
BDNF在脊髓各型神经无有明显的表达,特别是前角运动神经元。
trkA、trkB、trkC的免疫阳性反应产物主要分布在灰质的神经元及胶质细胞。
本实验结果揭示了在正常猴脊髓中神经营养因子(NGF、BDNF)及受体trkA、tr kB、trkC的表达状况,提示这些神经营养因子及受体在维持猴脊髓神经元的正常生理功能中具有重要作用。
关键词 神经营养因子 神经营养因子受体 免疫组织化学 脊髓 猴THE EXPRESSION OF NEUROTROPHIC FACTORS(NGF、BDNF)AND RECEPTORS, TrkA,TrkB AND TrkC IN THE SPINAL CORD OF THE MONKEYW u X iaoqiong,L i Changqi,L iu Dan,Cai W eij un,L uo X uegang(Department of Neurobiolog y,Xiang Ya M edical College,Central South University,Changsha410078)Abstract T o inves tigate the expression of neurotrophic factor s and their receptors in th e spinal cord of adult monk ey,im muno-histoch emis try w ith antibodies again st neurotrophic factors nerve grow th factor(NGF),brain derived neu rotr op hic factor (BDNF)and the neurotrophic factor r eceptors,trkA、trkB and trkC w as us ed.NGF immu noreactive positive neurons w ere pr e-sen t in all the laminae of g rey m atter,glial cells in the grey and wh ite matter w ere positively stained for NGF.BDNF immun or e-active pos itive neurons w ere obs erved in differen t neu rons of spinal cord,especial in m otoneurons,trkA、trkB and trkC im-munoreactivity w as obs erved in neurons and glial cells of the g rey matter.T he data suggest th at neu rotroph ins may play an im-portant role in maintain ing th e physiolog ical fun ction of neu rons in th e spinal cord.(Figu res a~h on plate50) Key words neurotrophic factor s,neurotroph ic receptors,immu nohistoch emis try,s pinal cord,monkey 神经生长因子(NGF)和脑源性神经营养因子(BDNF)属于神经营养因子家族的成员,它们对维持神经元的存活、生长、分化以及损伤后修复与再生有非常重要的作用[1]。
神经生长因子及受体在肿瘤中的表达及临床意义

神经生长因子及受体在肿瘤中的表达及临床意义神经生长因子(NGF)是一种多功能的神经营养因子,与特异的靶细胞表面的神经生长因子受体结合,介导产生一系列的生物学效应。
目前研究表明NGF 及其受体与肿瘤的增殖、分化、凋亡、血管形成、转移及预后也密不可分,且由NGF介导的信号通路已成为肿瘤治疗新的靶点。
标签:神经生长因子;受体;肿瘤神经生长因子(nerve growth factor,NGF)是一种多功能、多肽性的物质,具有刺激细胞生长活性的细胞因子。
一类通过与特异的、高亲和的细胞膜受体结合,调节细胞生长与其他细胞功能等多效应的多肽类物质。
它通过特异的靶细胞表面的神经生长因子受体(nerve growth factor receptor,NGFR)介导产生生物学效应。
近年来NGF及NG-FR与肿瘤的关系受到人们的关注。
本资料就神经生长因子及其受体与肿瘤的研究及进展作一综述。
1 神经生长因子神经生长因子是神经系统最重要的生物活性分子之一,是由其效应神经元支配的靶细胞合成和分泌,可以促进感觉神经元及交感神经元的存活及分化,参与损伤修复,维持其生物学功能的稳定。
NGF是由α、β、γ 3种亚基构成的多聚体,β亚基是唯一具有NGF的所有生物活性的亚基,现研究发现前列腺癌、乳腺癌和胰腺癌等肿瘤细胞也可合成NGF等细胞生长因子,并通过自分泌和旁分泌作用于周围的基质成分,来调节肿瘤的生长、分化和浸润行为,包括肿瘤的沿神经浸润。
NGF阳性表达随着肿瘤恶性程度的增高而增高,且与肿瘤的淋巴转移密切相关[1]。
2 NGF的受体(TrkA、p75)2.1 TrkATrkA是一种由原癌基因表达,具有酪氨酸激酶活性的跨膜蛋白,由跨膜酪氨酸激酶gp140Trk组成,是酪氨酸蛋白激酶家族成员之一。
由3个部分组成:(1)辨别并结合NGF的细胞外部;(2)跨膜部;(3)含酪氨酸激酶的胞质部。
TrkA称为高亲和力受体,也叫慢受体或I型受体[2]。
神经营养因子的两类受体:Trks及p75(综述)

中国神经免疫学和神经 苤圭
生 旦蔓 ! 塑 鲞笙
』!
! ! ! :
: !! : :!
神经 营养 因子 的两类受 体 : k Tr s及 p N ) (  ̄ 7 5
杨 卫 红 林 敬 明 ,
( 第一军医大学附属珠江 医院 ..神经内科 ,.药剂科 , 1 2 广东 广州 5 0 8 ) 1 2 2
Tr B结 合 。 k
13 T k . r s的激 活 Tr s包 括 Tr A、 k k
Tr k与 NT 结 合 后 形 成 二 聚
体 并 被 初 步 激 活一 其 “ 化 圈 ” 的酪 氨 酸 被 磷 酸 活 内
化, 继而 整 个 Tr k被 激 活 一 其 “ 化 圈 ” 的 酪 氨 活 外 酸 自动磷 酸 化 并 与 信 号 传 导 分 子 连 接 一 使 信 号 传
摘 要 :此文 简要 回顾了 神经营养 因子的两 类受体 —— 酪氨酸 激酶受体 家族 ( k ) p 5的分子 结构 、 布 、 Trs  ̄ 7 分 相 互作用 , 与神经营 养因子的结 合及 其后的信 号传导路径 。
关 键 词 :Trs 7 ;神 经 营 养 因 子 k ;p 5
中 图 分 类 号 :R3 8 文 献 标 识 码 :A 文 章 编 号 :1 0 —9 3 (0 2 0— 2 40 3 0 62 6 2 0 ) 40 4 —3
神 经 营 养 因子 ( NT) 族 包 括 神 经 生 长 因 子 家
( NGF 、 衍 生 的 神 经 营 养 因 子 ( D ) NT一 、 )脑 B NF 、 3 NT一 /  ̄ , 们 之 间 有很 高 的 同 源性 , 4 5I 他 ] 作用 也 很 相 近 , 受体 分 为 两 类 : 氨 酸 激 酶 受体 家 族 ( r s 其 酪 T k)
生长因子及其受体的作用机制研究

生长因子及其受体的作用机制研究生长因子和其受体是细胞生长和分化的重要信号通路。
生长因子是一类能够刺激细胞增殖和分化的蛋白质分子,其受体是一类跨越细胞膜的蛋白质,位于细胞膜上,可以接受外界刺激并转导信号到细胞内部。
1. 生长因子及其受体的种类目前所知的生长因子及其受体有很多种。
其中,最常见的包括:(1)表皮生长因子(Epidermal growth factor,EGF)及其受体(EGFR):EGF是一种重要的生长因子,可以刺激皮肤和黏膜上皮细胞增殖和分化。
EGFR受体是一种酪氨酸激酶受体,具有自身的酪氨酸激酶活性,参与调控细胞增殖、分化和凋亡等生命过程。
(2)神经生长因子(Nerve growth factor,NGF)及其受体(TrkA):NGF是神经生长和维持神经元生存的重要因子。
TrkA是NGF的高亲和性受体,调控神经细胞的生长、发育和再生。
(3)血小板衍生生长因子(Platelet-derived growth factor,PDGF)及其受体(PDGFR):PDGF是一种促进血管生成和组织再生的生长因子。
PDGFR是一种酪氨酸激酶受体,调控细胞增殖、迁移和血管生成等过程。
2. 生长因子及其受体的作用机制生长因子与其受体之间的信号转导机制包括以下几个方面。
(1)受体激活:生长因子与其受体结合后,会导致受体二聚化,即两个受体分子相互结合形成二聚体。
二聚化后的受体会包含自身酪氨酸激酶活性,从而使受体激活。
(2)下游途径:激活的生长因子受体会通过下游信号途径传递信号。
这些下游途径可以包括激酶级联反应、磷脂酰肌醇信号通路等多个信号通路。
(3)转录调控:下游信号途径会影响到细胞内的基因表达。
通过这种机制,生长因子能够调控细胞增殖、分化和凋亡等生命过程。
3. 生长因子及其受体在疾病中的作用生长因子及其受体在多种疾病中起到重要作用。
(1)癌症:生长因子及其受体在癌症的发生发展中起到至关重要的作用。
癌细胞的生长和转移需要某些生长因子和其受体的参与,在肿瘤组织中这些因子和受体的表达往往比正常细胞更高。
综述范文

医学免疫学综述泸州医学院学号姓名神经生长因子在哮喘中作用的研究进展【摘要】神经生长因子(nerve growth factor,NGF)[1]是神经元细胞生长和分化的营养因子,对神经细胞的生长、分化、存活、维持均起着重要作用。
NGF还通过调节多种免疫细胞,促进炎性介质释放,导致气道炎症,诱导神经元可塑性。
近来发现它不仅与炎性细胞的调节有关,参与炎症反应,而且NGF介导支气管哮喘(哮喘)时气道高反应性的形成,其水平在支气管哮喘患者血清、痰液、支气管肺泡灌洗液、支气管黏膜中均有明显升高。
因此,NGF在哮喘的相关研究中受到越来越多的重视。
[1]【关键词】神经生长因子;支气管哮喘;气道高反应性;气道炎症;气道重塑0 前言支气管哮喘(bronchial asthma)是有多种细胞特别是肥大细胞、嗜酸性粒细胞和T淋巴细胞参与的慢性气道炎症。
哮喘是一种常见病、多发病,发病率较高,近年对于哮喘的研究取得了很大的进展,但是哮喘的发病机制非常复杂,至今尚未完全阐明。
[2]近年来的研究发现在哮喘发病机制中,免疫细胞和神经元之间存在着广泛的联系,神经生长因子(NGF) 由免疫细胞产生,不仅作用于神经系统,它也影响机体免疫和造血系统,与变态反应性和炎症性疾病的发病有关,但是否参与了哮喘的病理生理过程以及它在哮喘的发病中起到何种作用,目前并不十分清楚。
一些学者认为NGF通过调控神经元的可塑性,介导气道高反应性,并参与气道炎症形成的过程,从而认为NGF是哮喘免疫神经发病机制的重要介质。
另有研究提示,NGF作为神经可塑性(neuronal plasticity)的调控因子,通过诱导神经源性炎症反应参与哮喘发病机制中。
本文就NGF的结构、功能调节以及与支气管哮喘的关系等方面作一综述。
1 NGF的结构NGF属于神经营养因子家族(neurophic factor,NTs),这是一组超出维持生存所必需的基本营养物质以外的,对神经细胞起特殊营养作用的多肽分子。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
神经生长因子受体TrkA在中枢神经系统的表达、分布和作用关键词: TrkA ChAT 基底前脑阿尔茨海默病雌激素摘要酪氨酸激酶(tyrosine kinase,TrkA)是神经生长因子(nerve growth factor,NGF)的功能性受体。
胆碱乙酰化转移酶(choline acetyltransferase,ChAT)是胆碱能神经递质乙酰胆碱合成的关键酶。
大鼠发育过程中基底前脑TrkA、ChAT表达有一定的规律,老龄鼠和早老性痴呆病人基底前脑TrkA、ChAT表达明显下调。
TrkA、ChAT雌激素受体共存于基底前脑神经元。
雌性激素对于大鼠基底前脑TrkA、ChAT维持正常的水平发挥作用。
雌性激素替代治疗绝经后的妇女有助于减少其患阿尔茨海默病的可能性。
关键词:TrkA ChAT 基底前脑阿尔茨海默病雌激素神经生长因子(nerve growth factor,NGF)是一种经典的神经营养因子。
NGF在周围神经系统(peripheral nervous system,PNS)参与交感神经元、神经嵴起源的感觉神经元的发育、存活,维持及损伤修复等作用已逐渐为人们所认识。
八十年以后,人们发现NGF对中枢神经系统(certral nervous system,CNS)中基底前脑(basal forebrain)胆碱能神经元有作用。
而作用于该部胆碱能神经元的NGF主要由靶区(海马及新皮质)产生,经逆行运输到胆碱能神经元胞体,发挥其靶源性营养作用。
大量的研究表明这类神经元亚群在学习和记忆中具有特殊功能。
在培养PC12细胞的培养基中加入NGF时,能诱导PC12突起的生长并使酶的活性提高,但当注射NGF到PC12细胞体或胞核时,并不发生以上变化,从而说明NGF生理功能的启动是由膜受体介导的。
进一步研究发现,NGF效应细胞膜上有两种受体类型,根据其对NGF的亲和性分为高亲和力受体(high affinity receptor,HNGFR)和低亲和力受体(low affinity receptor,LNGFR)。
LNGFR 是一种富含半胱氨酸的糖蛋白,其分子量为75kD,所以又称p75。
LNGFR胞质部分没有ATP结合位点,致使NGF与p75结合后不能活化内源性激酶,故NGF与p75的结合不能直接发挥生物学效应,但能通过增加HNGFR与NGF的结合率,影响通过HNGFR进行的信号传递。
1991年Klein[1]等研究发现HNGFR是由酪氨酸激酶原癌基因(tyrosine kinase proto-oncogene,TrK)表达的一种跨膜糖蛋白,分子量为140kD。
现已知HNGFR至少有一个亚基由TrkA原癌基因编码。
TrkA由三部分组成:即辨别并结合NGF的细胞外部、跨膜部及含酪氨酸激酶的胞质部。
TrkA是NGF 的功能性受体,当其与NGF结合后,可激活酪氨酸激酶信号传递系统,从而启动细胞活性,产生生物效应。
Boissiere[2]等用免疫组化方法检测人基底前脑,发现99%胆碱能神经元有TrkA表达;有人用原位杂交和免疫组化方法发现TrkA mRNA和蛋白质广泛地分布于大鼠中枢神经系统[3]。
在基底前脑表达TrkA mRNA的胆碱能纤维广泛地投射到海马和新皮质。
1 发育过程中基底前脑TrkA表达的变化利用免疫组化方法证实在成年大鼠基底前脑的胆碱能神经元有p75和TrkA的表达[4],有人用逆转录聚合酶链式反应(RT-PCR)探及到胚胎17天大鼠基底前脑胆碱能神经元无TrkA mRNA表达[5],生后3天到2周才发现有TrkA mRNA的表达;Li[6]等用原位杂交和Northern印迹等方法定量分析TrkA mRNA、ChAT mRNA 在大白鼠胚胎、生后、成年基底前脑内侧隔核(MS)的变化。
发现胚胎17天时均无表达,生后0天少数几个细胞表达TrkA mRNA、ChAT mRNA;生后4天至11天,两者表达明显增加,生后21天表达最强,高于成年鼠的水平。
生后30天呈下降趋势,在成年鼠维持在一个相对稳定比较高的水平。
TrkA mRNA和ChAT mRNA的表达有相似的时空模式。
最近有人用免疫组织化学方法观察TrkA表达,无ChAT表达,生后5天可见ChAT表达,生后20天TrkA mRNA表达达高峰;生后30天下调,成年时维持相对较高水平,老年时TrkA mRNA表达明显下调。
大鼠基底前脑Meynert 基底核TrkA表达早于ChAT表达。
从生后5天起,TrkA、ChAT表达有相似的时间模式。
TrkA可能参与Meynert基底核胆碱能神经元的发育、分化、成熟。
NGF mRNA、TrkA mRNA在发育过程中的变化对于基底前脑胆碱能神经元的存活及突起形成可能起一定的调节作用。
在脑发育不同阶段基底前脑胆碱能神经元TrkA基因表达具有阶段性差异,Hsiang[7]等用原位杂交方法发现,在出生后1天大鼠基底前脑胆碱能神经元没有测到TrkA基因,以后逐渐增加,4周时达最高水平,这种发育阶段的变化与海马及皮质中NGF mRNA基因表达的变化有平行关系[8],提示NGF 对胆碱能神经元发育的调控作用,也反应了这个过程中NGF受体对这种调控的参与。
2 老龄鼠和阿尔茨海默病(Alzheimer’s disease,AD)病人基底前脑胆碱能神经元TrkA、ChAT表达的变化Cooper[9]等将125I-NGF注射入成年大鼠和老龄大鼠(26~30个月)海马内,发现老龄鼠内侧隔核能摄取和逆行转动125I-NGF的ChAT免疫反应神经元数目减少31%,不能摄取和逆行转运125I-NGF的胆碱能神经元严重萎缩、胞体的平均面积减少60%,同时,TrkA mRNA表达下调43%,而p75没有差异。
表明老龄鼠基底前脑胆碱能神经元维持受体介导靶源性神经营养因子的摄取和逆行转运能力下降,导致NGF信号传递功能削弱。
增加了老年动物胆碱能神经元变性的易感性。
最近我们用免疫组织化学方法发现老龄鼠(Meynert)基底核(basal nucleus of Meynert,nBM)TrkA、ChAT-IR神经元萎缩、数量分别减少31.9%、37.5%;胞体平均截面积分别减少39.4%、30.4%;平均灰度分别减少11.8%、9.9%。
早老性痴呆是由于基底前脑胆碱能神经元的变性死亡及相应皮质、海马神经元退化所引起的老年神经元退行性疾病,其主要临床表现为学习及记忆功能障碍,导致严重的智力低下。
多数学者认为AD病因主要与乙酰胆碱能系统改变有关。
Boissiere[10,11]等用原位杂交方法发现AD病人与老年人对照组比较,nBM胆碱能神经元TrkA mRNA 表达减少75%,差异有显著性(P<0.001);腹侧纹状体(ventral atriatum)TrkA mRNA表达减少41%(P<0.01)。
豆状核壳(putamen)TrkA mRNA表达减少43%~53%(P<0.01)。
Mufson[12]等用原位杂交方法发现AD病人与老年人对照组比较,nBM胆碱能神经元TrkA mRNA表达减少66%,用逆转录聚合酶链式反应(RT-PCR)方法也得出同样的结论。
进一步用免疫组化方法研究发现:AD病人nBM内胆碱能神经元TrkA蛋白质表达与老年人对照组比较,AD病人nBM内幸存的TrkA iR 阳性神经元数量减少60%,染色密度减少35%,细胞外形皱缩,树突截断变形,整个细胞呈肿胀的球形外观(swollen globose appearance)[13]。
3 TrkA对于基底前脑胆碱能神经元的作用Fagan[14]等将小鼠基底前脑、纹状体胆碱能神经元编码TrkA受体的基因失活(TrkA-/-)发现生后7天小鼠基底前脑、纹状体胆碱能神经元ChAT-IR阳性产物胞体的平均截面积减少10%~20%;生后20~25天小鼠ChAT-IR阳性神经元数目20%~36%,胞体萎缩变小,ChAT免疫反应性降低,胞体和胆碱能神经纤维毡标记密度减少,表明正在发育的(TrkA-/-)小鼠胆碱能神经元ChAT表达减少,阻碍发育成熟。
结果表明正在发育(TrkA-/-)小鼠胆碱能神经元ChAT的表达减少,细胞死亡增加。
(TrkA-/-)小鼠基底前脑胆碱能神经元丢失与生后1~4周时,靶源性神经营养因子缺失导致细胞死亡是一致的[15]。
缺乏TrkA受体,NGF不能与特异性的受体结合,不能进行细胞内信号传递,正在发育的基底前脑胆碱能神经元不能充分地发育成熟。
4 TrkA与ChAT、雌激素受体(estrogen receptor,ER)共存Sobreviela[16]等用免疫组化方法发现:内侧隔核和斜角带核内95%以上TrkA 免疫反应阳性神经元内既含有ChAT也含有p75 nGFR;Meynert基底核内80%以上TrkA免疫反应阳性神经元内含有ChAT,95%以上的TrkA免疫反应阳性神经元内含有p75。
Gibbs[17]等用免疫组化方法证明50%~80%的胆碱能ChAT免疫反应神经元内含有ER,雌性大鼠较雄性大鼠多10.5%。
纹状体内双标细胞占74.2%,斜角带核水平支内为63.4%。
Toran[18,19]等用放射自显影和原位杂交、免疫组化方法发现基底前脑胆碱能神经元含有雌性激素的高亲和性连接位点,即雌激素受体,属核受体,为一核转录因子。
结果提示它们的配体(神经营养因子、雌性激素)可能作用于同一神经元,协同调节细胞内特异的基因或者基因网络的表达。
从而调控mRNA的细胞内组成,影响蛋白质生成的量及其性质,最终影响神经元的存活、分化、再生和可塑性。
5 雌性激素对于基底前脑胆碱能神经元TrkA mRNA和ChAT mRNA表达的调节Pamela[20]等人将成年大白鼠卵巢切除后10天,HDB和nBM内TrkA mRNA表达水平分别下调56%和34%,而ChAT mRNA分别下调38%~65%,与Gibbs[21]等人研究结果相似。
雌性激素替代治疗3天后,TrkA mRNA、ChAT mRNA表达恢复到未切卵巢的动物的水平,而VDB中的TrkA mRNA、ChAT mRNA表达没有明显的差异。
Gibbs[22]等观察鼠龄分别为13个月、19个月和25个月的雄性、雌性大鼠,发现年龄对于MS、nBM内ChAT、p75免疫反应阳性神经元的细胞大小、纤维染色密度的影响没有显著性差异,13~25个月龄鼠中,25个月龄鼠TrkA mRNA明显地减少。
将13个月龄鼠卵巢切除6个月后,MS、nBM内TrkA mRNA、ChAT mRNA明显地减少,短期内雌性激素替代治疗后,MS、nBM内TrkA mRNA、ChAT mRNA明显地减少,短期内雌性激素替代治疗后,MS、nBM内TrkA mRNA、ChAT mRNA部分恢复。