第三章 免疫球蛋白和抗体 1
合集下载
三免疫球蛋白与抗体【40页】

免疫球蛋白的生物学功能
四、通过胎盘和黏膜 IgG是人类唯一能够从母体通过胎盘屏障转运到胎儿体内的抗体。IgG通过 其Fc段选择性地与胎盘微血管壁内皮细胞表面受体可逆性结合,主动穿过 胎盘进入胎儿血液循环。 此外,SlgA可通过分泌片经黏膜上皮细胞进入呼吸道及消化道发挥抗感染 免疫作用。
目录
各类免疫球蛋白的特性和作用
免疫球蛋白的生物学功能
二、激活补体 lgM、IgG抗体与相应抗原结合后,抗体的构象改变使其CH2或CH3区的补体 C1q结合位点暴露,从而导致补体经典途径激活。凝聚的gA1、lgG4和IgE可 以通过旁路途径活化补体。
免疫球蛋白的生物学功能
三、结合Fc受体 lgG、lgA和IgE可通过其Fc段与细胞表面的相应Fc受体(FcγR、FcαR、FcεR)结 合,发挥免疫效应。 1.调理作用指抗体通过Fab段与细菌等颗粒性抗原结合,以Fc段结合巨噬细 胞或中性粒细胞表面的FcγR,从而促进吞噬细胞对上述颗粒性抗原吞噬的 作用。
人工制备抗体
2.单克隆抗体由单一B细胞克隆产生的、只识别单一抗原表位的高度特异性 抗体称为单克隆抗体( monoclonal antibody,mAb)。以免疫小鼠的B细胞与同 系小鼠骨髓瘤细胞融合形成的杂交瘤( hybridoma)细胞,既有骨髓瘤细胞无限 扩增的特性,又继承了B细胞合成特异性抗体的能力。一个杂交瘤细胞增殖 形成的细胞群所产生的抗体是只能识别一种抗原表位的mAb。mAb的优点是 结构均一、纯度高、特异性强,几乎不存在交叉反应,目前已在医学、生物 学各领域广泛应用。 3.基因工程抗体通过基因重组技术制备的抗体称为基因工程抗体( genetic engineering antibody)。这种抗体既保留了单克隆抗体的均一性、特异性强的 优点,又能克服其鼠源性的缺点。如人鼠嵌合抗体、改型抗体及小分子抗体 等。
4-1第三章第三节免疫球蛋白与抗体

单克隆抗体制备示意图
PEG:聚乙二醇
三、基因工程抗体
五类免疫球蛋白
二、免疫球蛋白的结构和功能
• 3.免疫球蛋白的水解片段
二、免疫球蛋白的结构和功能
(二)生物学功能(特异 性结合抗原)
• • • • 1.Fab段 (1)结合的部位:V区 (2)结合有特异性 (3)结合的力学特点:氢键、 静电、范德化力 • (4)可逆性 • (5)影响因素:pH值、温 度、电解质等 • (6)价位
二、免疫球蛋白的结构和功能
• ②抗体依赖的细胞介导的细胞毒作用(ADCC)
• antibody-dependent cell-mediated cytotoxicity
二、免疫球蛋白的结构和功能
• ③介导Ⅰ型变态反应
二、免疫球蛋白的结构和功能
• (3)穿过胎盘与黏膜(IgG、SIgA)
三、各类免疫球蛋白的特性和功能
第三节 免疫球蛋白
第三节 免疫球蛋白
• 抗体(antibody,Ab)是B细胞接受抗原刺激后 增殖分化为浆细胞,由浆细胞产生的一类能与相 应抗原特异性结合的球蛋白。 • 免疫球蛋白(immunoglobulin,Ig):具有抗体 活性或化学结构与抗体相似的球蛋白。 • 抗体都是免疫球蛋白,免疫球蛋白不一定都具有 抗体活性。 • Ig分泌型:存在血液、组织液中,抗体功能 • Ig膜型:构成B细胞膜上的抗原受体。
• • • •
2.Fc段: (1)激活补体 (2)结合细胞 (3)穿过胎盘和黏膜
二、免疫球蛋白的结构和功能
• • • • • (1)激活补体 能激活补体的免疫球蛋白: IgG1~IgG3、IgM的CH区——补体C1q IgA1、IgG4旁路 IgD、IgE不能
二、免疫球蛋白的结构和功能
PEG:聚乙二醇
三、基因工程抗体
五类免疫球蛋白
二、免疫球蛋白的结构和功能
• 3.免疫球蛋白的水解片段
二、免疫球蛋白的结构和功能
(二)生物学功能(特异 性结合抗原)
• • • • 1.Fab段 (1)结合的部位:V区 (2)结合有特异性 (3)结合的力学特点:氢键、 静电、范德化力 • (4)可逆性 • (5)影响因素:pH值、温 度、电解质等 • (6)价位
二、免疫球蛋白的结构和功能
• ②抗体依赖的细胞介导的细胞毒作用(ADCC)
• antibody-dependent cell-mediated cytotoxicity
二、免疫球蛋白的结构和功能
• ③介导Ⅰ型变态反应
二、免疫球蛋白的结构和功能
• (3)穿过胎盘与黏膜(IgG、SIgA)
三、各类免疫球蛋白的特性和功能
第三节 免疫球蛋白
第三节 免疫球蛋白
• 抗体(antibody,Ab)是B细胞接受抗原刺激后 增殖分化为浆细胞,由浆细胞产生的一类能与相 应抗原特异性结合的球蛋白。 • 免疫球蛋白(immunoglobulin,Ig):具有抗体 活性或化学结构与抗体相似的球蛋白。 • 抗体都是免疫球蛋白,免疫球蛋白不一定都具有 抗体活性。 • Ig分泌型:存在血液、组织液中,抗体功能 • Ig膜型:构成B细胞膜上的抗原受体。
• • • •
2.Fc段: (1)激活补体 (2)结合细胞 (3)穿过胎盘和黏膜
二、免疫球蛋白的结构和功能
• • • • • (1)激活补体 能激活补体的免疫球蛋白: IgG1~IgG3、IgM的CH区——补体C1q IgA1、IgG4旁路 IgD、IgE不能
二、免疫球蛋白的结构和功能
第三章-免疫球蛋白和抗体(a)课件

❖同一种属不同个体间,同一类型的Ig分 子所具有的不同的抗原特异性标志,为 遗传标志,存在于重链或轻链恒定区内, 表现为一个或数个氨基酸的差异。
35
36
独特型(idiotype,Id)
❖每个Ig分子可变区肽链所具有的抗原特 异性标志——Id。独特型表位又称独特 位,是由Ig超变区特定序列和构象的氨 基酸所组成。独特位在异种、同种异体 和同一体内均有可能刺激机体产生抗独 特型抗体(AId)。
❖同种型 ❖同种异型 ❖独特型
32
同种型(isotype)
❖指同一种属所有正常个体同一类型Ig分 子所共有的抗原特异性标志,主要存在 于Ig的CH和CL中。
❖据H链C区的不同又分为五个大类,每一 类又分若干亚类;按L链C区的不同分为 两型,每型又分为若干亚型。
33
34
同种异型(allotype)
及两条相同的轻链 以-S-S-相连的四肽 链结构。
6
重链和轻链 ❖重链(H链)
➢ 50-70KD,450-550个氨基酸组成 ➢ 分成五类,相应Ig也分为五类。 H: α γ μ δ ε Ig: IgA IgG IgM IgD IgE
8
➢亚类—— 同一类Ig据其铰链区的氨基酸组成及重链二硫键 数目将Ig分为不同的亚类。
16
17
❖ 铰链区—— 位于CH1与CH2之间,含有丰富的脯氨
酸,因此易伸展弯曲,而且易被木瓜蛋白 酶、胃蛋白酶等水解。
18
19
(二)Ig功能区的结构和主要功能
❖结构——Ig折叠 Ig的二级结构是由几股多肽链折叠形成 的两个反向平行的β片层,两个β片层中 心由链内二硫键垂直连接,形成“β桶状” 或“β三明治”结构。这种折叠方式称为免 疫球蛋白折叠。
35
36
独特型(idiotype,Id)
❖每个Ig分子可变区肽链所具有的抗原特 异性标志——Id。独特型表位又称独特 位,是由Ig超变区特定序列和构象的氨 基酸所组成。独特位在异种、同种异体 和同一体内均有可能刺激机体产生抗独 特型抗体(AId)。
❖同种型 ❖同种异型 ❖独特型
32
同种型(isotype)
❖指同一种属所有正常个体同一类型Ig分 子所共有的抗原特异性标志,主要存在 于Ig的CH和CL中。
❖据H链C区的不同又分为五个大类,每一 类又分若干亚类;按L链C区的不同分为 两型,每型又分为若干亚型。
33
34
同种异型(allotype)
及两条相同的轻链 以-S-S-相连的四肽 链结构。
6
重链和轻链 ❖重链(H链)
➢ 50-70KD,450-550个氨基酸组成 ➢ 分成五类,相应Ig也分为五类。 H: α γ μ δ ε Ig: IgA IgG IgM IgD IgE
8
➢亚类—— 同一类Ig据其铰链区的氨基酸组成及重链二硫键 数目将Ig分为不同的亚类。
16
17
❖ 铰链区—— 位于CH1与CH2之间,含有丰富的脯氨
酸,因此易伸展弯曲,而且易被木瓜蛋白 酶、胃蛋白酶等水解。
18
19
(二)Ig功能区的结构和主要功能
❖结构——Ig折叠 Ig的二级结构是由几股多肽链折叠形成 的两个反向平行的β片层,两个β片层中 心由链内二硫键垂直连接,形成“β桶状” 或“β三明治”结构。这种折叠方式称为免 疫球蛋白折叠。
第3章 免疫球蛋白

三、免疫球蛋白的水解片断
1、水解作用
Ig 木瓜蛋白酶 2Fab+Fc Ig 胃蛋白酶 F(ab´)2+pFc´
2、 水解意义
阐明Ig的结构和生物活性, 指导生物制品的生产和临床应用
第二节 五类免疫球蛋白 的特点和功能
一、Ig分类 1、类:根据同种系所有个体的Ig重链C区的抗原性 分为五类 IgM()、IgG()、IgA()、IgD()、IgE() 2、亚类:同一类Ig内的aa序列及二硫键的数目和位 置 IgG—IgG1、IgG2、IgG3和IgG4 ; IgA—IgA1和IgA2; IgM—IgM1和IgM2。 3、型:根据Ig轻链C区的抗原性分为链和链。 :约2:1. 4、亚型: 链有多种亚型。(轻链抗原决定簇在CL区, 分κ 和λ 。1个Ig分子轻链只有一个型,不是κ 型就 是λ 型。)
10,530
320 X 10,530 = 3,369,600
The potential diversity generated by V(D)J recombination is virtually infinite. >Total lymphocytes in a human ~1012 >Genome size ~3X109 bp
四、选择性传递
(主要借助胎盘滋养层细胞表面的FcR进行的。 )
五、具有免疫原性
1、同种型(isotype)
2、同种异型(allotype)
3、独特型(idiotype)
将Ig作为抗原在异种、同种异体或自身体 内进行免疫,均能诱导机体发生免疫应答, 产生相应的抗体,这些抗体可用血清学方 法进行检测,因此称为Ig的血清型
(二)重链类型转换 (成熟B细胞,抗原刺激后) 即class switch,又称S-S转换。
第三章 免疫球蛋白与抗体

种类型的重链, γ 、μ、α、 ε、 δ链,分别对应IgG,
IgM,IgA, IgE和IgD。 (2)轻链(light chain,L链):由213- 214个氨基酸残 基组成,分子量为22.5KU,氨基端约1/2部分为轻链可变 区。根据L链稳定区氨基酸序列的不同, L链可分为: Kappa(κ)与lambda(λ)2个亚型。 不同的H链与L链组成完整的Ig分子。
除轻、重链V区以外的部分分别为轻、重链的C区。
(三)免疫球蛋白的功能区(domain)
免疫球蛋白的多肽链分子折叠形成几个由链内及链间二硫键形成的环状球 性结构,称为免疫球蛋白的功能区。每一个功能区都约有110个氨基酸组成
VH-VL高变区为抗体与抗原的结合的部位,称为抗体分子与抗原决定簇的
互补决定区(complementarity-determining region, CDR)。该区域决定了抗 体分子的特异性。 CL-CHI: 遗传标志所在。 CH2:抗体分子的补体结合位点。 CH3:与抗体分子的亲细胞性有关,是Ig同一些免疫细胞的Fc受体结合的
同种异体和同一个体内均可产生相应的抗体,这种抗体成为抗独特型 抗体。
片灯 幻 张 一 第
第4节 各类免疫球蛋白的主要特性与免疫学功能
各类免疫球蛋白的主要特性与免疫学功能见p47。 (1)IgG是人和动物血清中含量最高的免疫球蛋白。占血
清免疫球蛋白的75%-80%。
(2)IgM是动物机体初次体液免疫反应最早产生的免疫球 蛋白。占血清免疫球蛋白的10%左右。
第三章 免疫球蛋白与抗体
第1节 免疫球蛋白与抗体的概念
第2节 免疫球蛋白的分子结构
第3节 免疫蛋白的种类与抗原决定簇
第4节 各类免疫球蛋白的主要特性与免疫学功能
免疫学第三章抗体
❖ 同种型特异性因种而 异,如人类有五类 Ig、大鼠有七类Ig、 家兔有四类 Ig。
IgG1 (kappa)
IgM (kappa)
(2)同种异型(allotype)
❖ 同种系的不同个体之 间,同一类、亚类、型、 亚型的免疫球蛋白, 在结构上可能会有一 个或数个氨基酸的差 异, 称为同种异型。
❖ 由同一基因座位上不 同等位基因编码。
排列顺序以及结构是随 抗体分子的特异性不同而 有所变化,决定了抗体的 特异性。
重链的可变区和恒定区
❖ 恒定区(constant region, C区):可变区外其余 的氨基酸分子。(占重 链的3/4)。
数量、种类、排列 顺序及含糖量都比较 稳定,称为稳(恒)定 区。
轻链的可变区和恒定区
❖ 可变区(V区):氨基
❖ 免疫球蛋白的存在形式:
免疫球蛋白除分布于体液中之外,还可 存在于B细胞膜上。
❖ 膜型(membrane immunoglobulin, mIg)
--- B细胞膜上的抗原受体;
❖ 分泌型(secreted immunoglobulin, sIg)
---分泌进入体液, 介导体液免疫应答。
2、免疫球蛋白的基本结构
即抗原结合片段。
Fc 片段(fragment crystallizable,Fc):在
低温或低离子强度下可形成结晶,故称为可 结晶片段。
❖ Fab 片段:
每一个含有一条 完整的 L 链和 H 链近 N 端侧 的 1/2 。
❖ Fc 片段:
含有两条 H 链 羧基端(C 端) 的一半。
❖ Fab 片段:
❖ 五 类 免 疫 球 蛋 白 IgG , IgM , IgA , IgE , IgD的单体分子结构都是相似的,即是由 两条相同 H 链和两条相同 L 链共四条肽 链构成的“Y”字形的分子。
IgG1 (kappa)
IgM (kappa)
(2)同种异型(allotype)
❖ 同种系的不同个体之 间,同一类、亚类、型、 亚型的免疫球蛋白, 在结构上可能会有一 个或数个氨基酸的差 异, 称为同种异型。
❖ 由同一基因座位上不 同等位基因编码。
排列顺序以及结构是随 抗体分子的特异性不同而 有所变化,决定了抗体的 特异性。
重链的可变区和恒定区
❖ 恒定区(constant region, C区):可变区外其余 的氨基酸分子。(占重 链的3/4)。
数量、种类、排列 顺序及含糖量都比较 稳定,称为稳(恒)定 区。
轻链的可变区和恒定区
❖ 可变区(V区):氨基
❖ 免疫球蛋白的存在形式:
免疫球蛋白除分布于体液中之外,还可 存在于B细胞膜上。
❖ 膜型(membrane immunoglobulin, mIg)
--- B细胞膜上的抗原受体;
❖ 分泌型(secreted immunoglobulin, sIg)
---分泌进入体液, 介导体液免疫应答。
2、免疫球蛋白的基本结构
即抗原结合片段。
Fc 片段(fragment crystallizable,Fc):在
低温或低离子强度下可形成结晶,故称为可 结晶片段。
❖ Fab 片段:
每一个含有一条 完整的 L 链和 H 链近 N 端侧 的 1/2 。
❖ Fc 片段:
含有两条 H 链 羧基端(C 端) 的一半。
❖ Fab 片段:
❖ 五 类 免 疫 球 蛋 白 IgG , IgM , IgA , IgE , IgD的单体分子结构都是相似的,即是由 两条相同 H 链和两条相同 L 链共四条肽 链构成的“Y”字形的分子。
微生物兽医免疫第三章抗体与免疫球蛋白

2、胃蛋白酶 水解 IgG,获得一个F(ab′)2片段及若
干小分子片段,被称为pFc′。 (1)F(ab′)2片断:
为双价; (2)pFc′:
失去生物学活性 。
三、J链和分泌成分
1、J链(joining chain) (1)是一条多肽链,由浆细胞产生; (2)可连接Ig单体形成二聚体、五聚体。
IgG 440,IgM、IgE 550个氨基酸
(3)V区有3个区域的氨基酸组成和排列顺序 特别易变化,称为高变区;高变区之外区域 称为骨架区; (4)据重链 C 区氨基酸的组成和排列的不同, 将Ig分为五类。
IgG、IgM、IgA、IgD、IgE
2、轻链(light chain,L链) (1)由220个氨基酸组成; (2)轻链N端的109个氨基酸为V区,其余 为C区; (3)V区有3个高变区; (4)轻链分为κ(Kappa)和λ(Lambda) 两型。
猪瘟病毒 —— 抗猪瘟病毒抗体(猪) 口蹄疫病毒 —— 抗口蹄疫病毒抗体(猪) 猪丹毒杆菌 —— 抗丹毒杆菌抗体(猪) 猪免疫球蛋白G 免疫家兔 兔产生抗猪 免疫球蛋白抗体(抗抗体)
抗原 抗体(猪,IgG) 抗抗体(兔)
抗原
抗体
(猪瘟病毒,口蹄疫病毒,猪丹毒杆菌)
第七节 人工制备抗体的种类
多克隆抗体 : 多个抗原决定蔟——机体——多种
3、铰链区 (1)位于CH1与
CH2之间; (2)易于弯曲;
含有丰富的脯氨酸 (3)易被木瓜蛋白酶、
胃蛋白酶水解。
二、免疫球蛋白的水解片段及生物学活性 1、木瓜蛋白酶水解片段 Ig被木瓜蛋白酶水解后得到3个片段,2个相
同的 Fab 片段和1个 Fc 片段。
(1)Fab片段:即抗原结合片断 (antigen-binding fragment)
免疫球蛋白和抗体

学习目标
(2课时)
• 掌握:抗体和免疫球蛋白的概念; 免疫球蛋白的结构和功能;各类免 疫球蛋白的特性和功能。 • 熟悉:免疫球蛋白的水解片段;多 克隆抗体和单克隆抗体的概念。
• 了解:免疫球蛋白的异质性及抗体 的制备方法。
抗体的发现和研究历史:
1889年:绿脓杆菌
感染
动物血清
绿脓杆菌 1890年(Behring): 白喉类毒素
VL
V区的高变区(CDR)
CH1
CL
铰链区
CH2 CH3
CDR2(H)
CDR3(L)
CDR1(L)
抗原决定基
CDR2(L)
CDR1(H)
CDR3(H)
互补决定区和抗原决定基
恒定区(constant region, C区)
C端重链3/4,和轻 链1/2 组成(CH\CL) 氨基酸组成、序列稳定 某些抗体的C区具有补 体结合位点,具有多 种重要生物学活性
IgV区的功能:
与相应抗原特异性结合
Ig C区的功能:
激活补体 与细胞表面Fc受体结合 通过胎盘和黏膜
一、特异性结合抗原
由IgV区来实现 CDR 识别和结合特 异性抗原
形成Ag-Ab复合物, 使抗原不能发 挥原有的生物学效应
Toxin receptors
结果
Bacterial toxins
Host cell
中 和 作 用 通 过 胎 盘
结 合 受 体
超 用 敏 反 作 应 ADCC
体 外 反 应
课堂讨论
问题1:病毒感染是影响人类健康的重大疾病, 通过本节课的学习,Ab在抗病毒感染的过程作用 有哪些作用? 问题2:除了传统意义上的作用,你能否想象一 下,Ab是否有新的抗病毒感染的机制? 问题3:Ab抗病毒感染新机制对病毒感染控制有 什么提示?
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
• 3、抗体依赖性细胞介导的细胞毒作用 (ADCC) • IgG类抗体与肿瘤或病毒感染细胞表面相 应抗原表位特异性 结合后,再通过其Fc 段与NK细胞、巨噬细胞和中性粒细胞表 面相应IgGFc受体结合,增强或触发上述 效应细胞对靶细胞杀伤破坏的作用。
• 4、介导I型超敏反应:IgE为亲细胞性
• 2、IgM:分为膜结合型和血清型两种 类型,膜结合型IgM(mIgM)为单体, 表达于B细胞表面,构成B细胞抗原受 体,血清中IgM是由五个单体IgM通过 二硫键和连接链(J链)相连组成的五 聚体,相对分子质量居五类免疫球蛋 白之首,又称巨球蛋白。
• ①IgM不能通过血管壁,主要存在于血 液中,具有高效抗感染免疫作用; • IgM是个体发育过程中最早产生的抗 体,可在胚胎晚期生成,脐带血IgM含 量升高,提示胎儿宫内感染; • IgM是初次体液免疫应答中最早产生 的抗体
• 3、IgA:分为血清型和分泌型两种类 型,血清型IgA主要为单体IgA,分泌 型IgA(SIgA)是由J链连接的IgA二 聚体与一个分泌片借二硫键共价结合 组成。
• SIgA主要存在于呼吸道、消化道、泌 尿生殖道黏膜表面,以及乳汁、唾液 和泪液等外分泌液中。它们能与存在 黏膜局部的病原微生物结合,使之丧 失与细胞粘附的能力,从而在黏膜局 部发挥重要的抗感染免疫作用。
第三章 免疫球蛋白和抗体
• 一、教学目标 • 掌握免疫球蛋白和抗体的概念、免疫 球蛋白的功能、酶解片段及其作用 • 熟悉免疫球蛋白基本结构、类型和主 要生物学功能;各类免疫球蛋白的主 要特性和功能 • 了解人Ig胚系基因结构及其重排和免疫 球蛋白类别转换的感念
第一节 概述 • 1、抗体(Ab):是B细胞接受抗 原刺激,增殖分化为浆细胞后所产 生的一类具有免疫功能的糖蛋白。 主要存在于血清和其他体液中,是 介导体液免疫的重要效应分子。
• 二、免疫球蛋白恒定区(C区)的功 能 • 1、激活补体系统:IgG1-3和IgM与相 应抗原结合后,可因构象改变使其位 于CH2和CH3功能区内的补体C1q结 合位点暴露,从而导致补体经典途径 激活。
• 2、调理作用:IgG类抗体与相应细菌 等颗粒性抗原特异性结合后,通过其 Fc段与巨噬细胞或中性粒细胞表面相 应IgGFc受体结合,所产生的促进吞 噬细胞对上述颗粒性抗原吞噬的作用 称为调理作用。
• 1、重链:免疫球蛋白重链由 450~550个氨基酸残基组成。可分 为u、r、a、б、ε链,与轻链共同 组成Ig分别称为IgM、IgG、IgA、 IgD和IgE。 • 轻链:由214个氨基酸残基组成, 根据结构和抗原性的不同,分为k 和r两型。
五类免疫球蛋白:
• • • • • IgG 的H链:γ链 (gamma) IgA 的H链:α链 (alpha) IgM 的H链:μ链 (mu) IgD 的H链:δ链 (delta) IgE 的H链:ε链 (epsilon)
• u和ε重链恒定区除有上述三功能 区外,还有一个CH4功能区 • 轻链恒定区内只有一个功能区, 以CL表示。
• 3、超变区和骨架区 • 超变区:重链和轻链可变区结构域中, 三个特定区段内的氨基酸组成和排列 顺序有更大的变异性,这些区段称为 超变区(HVR),分别用HVR1, HVR2,HVR3表示。 • 骨架区:可变区中超变区之外的氨基酸 组成和排列顺序变化小,,称为骨架 区(FR)。
SIgA形成和转运
• ※新生儿易患呼吸道、消化道感染
性疾病,通过母乳可使新生儿从乳 汁中被动获得抗感染所需的SIgA, 因此应大力提倡母乳喂养。
• 4、IgD:分为血清型和膜结合型, 二者均以单体形式存在。前者的生 物学功能目前还不明确,其含量极 低。后者是表达于B细胞表面的抗 原受体,也是B细胞发育分化成熟 的标志。
四、免疫球蛋白的水解片段
1. 经木瓜蛋白酶水解IgG,得到3个片段, 2个相同的Fab段和1个Fc段
Fab段—抗原结合片段(fragment antigen binding), 每个Fab段由一条 完整的L链和一条约1/2 的H链组成, 具有抗体活性,能和一个抗原结合 表现为单价。
• Fc段— 可结晶片段(fragment crystallizable)由二条羧基端1/2H链组成, 不能和抗原结合,但有Ig的其他活性。 • 2 .经胃蛋白酶水解IgG,得到2个片段, • F(ab,) 2—具有双价抗体活性片段 • pFc’——小分子多肽碎片,无任何生物活 性
4、互补结合区和铰链区:重链和轻链可 变区结构域内三个超变区共同组成Ig的 抗原结合部位,该部位能与相应抗原 决定基互补结合,因此超变区又被称 为互补决定区(CDR)。不同抗体的 CDR序列不同,并因此决定了抗体与 相应抗原表位结合的特异性。
• Ig铰链区:位于重链CH1、CH2之间的 区域上,该区含有大量的脯氨酸,富 有弹性,易伸展弯曲,可使免疫球蛋 白分子由T型变为Y型,使其适合与抗 原分子表面不同距离的抗原表位,或 能同时与两个抗原分子表面相应的抗 原表位结合。(IgG、IgA、IgD重链
CH1与CH2之间有铰链区,IgM、 IgE重链无铰链区)
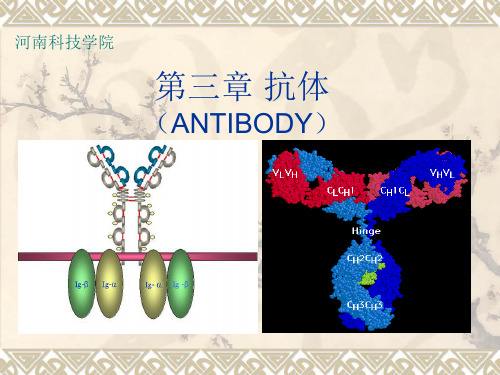
二、免疫球蛋白功能区结构和主要功能
• VH和VL共同构成抗原结合的部位。 • CL和CH1为同种异型遗传标志所在的部位。 • IgG的CH2和IgM 的CH3区是补体结合点所 在的部位,参与活化补体。 • IgG的CH2可介导IgG通过胎盘 • IgG的CH3和IgE的CH2、 CH3区能与组织 细胞表面的Fc受体结合。
抗体,可通过其Fc段与肥大细胞和嗜 碱性粒细胞表面相应IgEFc受体结合, 而使上述细胞处于致敏状态。致敏细 胞通过表面特异性IgE抗体与相应抗原 结合后,可脱颗粒、释放生物活性介 质,引起I型超敏反应。
• 5、穿过胎盘屏障和黏膜:人类IgG 是唯一能够从母体通过胎盘运转到 胎儿体内的免疫球蛋白。(分泌型 IgA可通过分泌片介导穿越呼吸道、 消化道等黏膜上皮细胞,到达黏膜 表面发挥抗感染作用)
抗原 非分泌性小鼠 骨髓瘤细胞 脾细胞
在PFG作用下细胞融合
转移到HAT培养基
骨髓瘤细胞被HAT杀死,脾细胞逐渐死亡, 融合细胞存活 杂交瘤细胞的筛选和克隆建株
筛选出能产生特异性抗体的杂交瘤细胞
单克隆抗体的制备示意图
三、J链和分泌片
• 1、J链:是一条富含半胱氨酸的多肽 链,由浆细胞合成,其主要功能是将 单体Ig分子连接成为多聚体。(IgG、 IgD、IgE和血清型IgA为单体分子,不 含J链)
• 分泌片(SP):又称分泌成分 (SC)是一种含糖的肽链,由黏 膜上皮细胞合成分泌。分泌片的主 要生物学作用:介导IgA二聚体 从黏膜下转运至黏膜表面;保护 分泌型IgA不被蛋白酶水解。
三、各类免疫球蛋白的主要特性和功能
• 1、IgG:主要存在于血液和组织夜中, 是人体内五类免疫球蛋白中含量最多 (75%~80%),是再次体液免疫应答 产生的主要抗体。是唯一能够通过胎 盘的抗体,在新生儿抗感染中起重要 作用。
• 功能:①IgG1-3与相应抗原结合形成 免疫复合物可激活补体经典途径; • ②IgG4凝聚物可激活补体旁路 途径; • ③IgG具有亲细胞特性,可通过 其Fc段与表面具有相应受体的吞噬细 胞和NK细胞结合,从而产生促进吞噬 的调理作用和ADCC效应。
Hale Waihona Puke 、免疫球蛋白的类型• 根据Ig重链恒定区肽链抗原特异性的不同, 可将免疫球蛋白分为IgM、IgG、IgA、 IgD和IgE。 • 现已发现:IgG有IgG1、IgG2、IgG3和 IgG4四个亚型;IgA有IgA1和IgA2亚型。
第三节 免疫球蛋白的生物学特性
• 一、免疫球蛋白可变区(V区)主要功能 • 免疫球蛋白可变区的主要功能是特异性识 别结合抗原,其内的超变区是与抗原表位 互补结合的区域。
• 5、IgE:是正常人血清中含量最低的 Ig,但在过敏性疾病或寄生虫感染患 者血清中,特异性IgE含量显著增高。 IgE为亲细胞性抗体,可通过其Fc段 与肥大细胞和嗜碱性粒细胞表面相应 IgEFc受体结合,而使上述细胞致敏。 由此导致I型超敏反应。
第四节 多克隆抗体和单克隆抗体
一、多克隆抗体(Polyclonal antibody)
用传统的抗体制备方法,将抗原物质注 入动物体内获得的免疫血清,是机体多个B 细胞克隆针对抗原分子上的不同抗原决定 簇产生的混合抗体,这种用体内免疫法制备 的免疫血清称为多克隆抗体.
• 二、单克隆抗体(monoclonal Ab,McAb) • •
概念: 由B细胞杂交瘤细胞株针对一 个抗原决定簇产生的纯抗体称单克隆抗体。 特点:纯度高、特异性强、效价高、 很少有交叉反应。已广泛用于生命科学的 各个领域。如:作为诊断试剂用于许多血 清学检测,用于治疗肿瘤,自身免疫性疾 病,抑制同种异体移植排斥反应。
• 1、免疫球蛋白(Ig):是指具有 抗体活性或化学结构与抗体相似的 糖蛋白。在血清中主要以r球蛋白 的形式存在。免疫球蛋白有分泌型 和膜型两种类型:前者主要存在于 血液和其他体液。后者作为抗原识 别受体表达于B细胞膜表面,称为 膜表面免疫球蛋白。
第二节 免疫球蛋白的结构 • 免疫球蛋白的基本结构(即Ig单体) 是由两条相同的重链(H链)和两 条相同的轻链(L链)通过键间二 硫键连接组成的一个四肽分子。
• 2、可变区与恒定区 • 可变区:免疫球蛋白重链近氨基端 的1/4或1/5区段内和轻链近氨基端 1/2区段,约110个氨基酸残基组成 和排列顺序多变,称为可变区(V 区) • 恒定区:羧基端轻链的1/2和重链 的3/4的区域内的氨基酸残基组成 和排列顺利相对稳定,称为恒定区。
• 功能区:重链和轻链可变区肽键通 过二硫键连接折叠,形成一个球状 结构域,又称为功能区。重链和轻 链可变区球形结构域分别以VH和 VL表示;重链和轻链恒定区可形 成几个球状结构域:r、a、б重 链恒定区内形成三个功能区→CH1、 CH2、CH3