实验二重组质粒酶切鉴定2016
重组质粒酶切与PCR验证

重组质粒的酶切和PCR验证1、学习和利用限制性内切酶酶切检验重组质粒插入片段的方法;2、复习限制性内切酶酶切原理和方法;3、学习和掌握PCR反应的基本原理和实验技术;4、学习利用PCR方法检验重组质粒插入片段的方法;5、了解引物设计的基本要求;【实验原理】1、重组质粒是外源基因通过单酶切与用相同的限制性核酸内切酶切割的质粒载体连接后获得的,因此在连接处仍保留原限制性核酸内切酶的识别序列,用该酶再次切割重组质粒,能把插入片段切离载体,根据琼脂糖凝胶电泳检测可见有载体片段和插入的外源DNA片段,并可知插入片段的大小。
2、聚合酶链式反应(Polymerase chain reaction),即PCR技术是一种体外快速扩增特定基因或DNA序列的方法,故又称为基因的体外扩增法。
PCR的反应组分包括:模板DNA、寡核苷酸引物、Mg2+、4中脱氧核糖核苷酸(dNTPs)、DNA聚合酶和适合的缓冲体系等。
反应包括DNA变性,引物复性和引物延伸三个基本过程。
①DNA变性,即在90-95℃高温下,加热使模板DNA双链解离,形成两条单链DNA;②引物复性,又称为引物退火。
将反应温度降低,使两个引物分别与变性的单链靶DNA特定部位结合形成部分双链DNA;③引物延伸,即在60-72℃以及四种脱氧核糖核苷三磷酸存在的条件下,由DNA 聚合酶催化,以单链靶DNA为模板,根据碱基配对原色,在引物3’端掺入dNTP合成新的DNA互补链,产生新的双链DNA。
3、PCR反应有两个重要特征:一是经PCR扩增后,扩增产物片段在大小由两个引物与模板的结合位点决定;二是经过PCR扩增,产物以指数级数增加,可达到目的片段的大量扩增。
在此实验中PCR技术实际上是在模板DNA、引物和4种脱氧核糖核苷酸存在的条件下依赖于DNA聚合酶的酶促合成反应。
PCR 技术的特异性取决于引物和模板DNA结合的特异性。
4、寡核苷酸引物或称PCR引物,是指两段与待扩增的目的DNA序列两侧翼片段互补的一段单链寡核苷酸序列。
重组质粒DNA的提取、 酶切鉴定

1.0 ml
+ 200l
悬浮液
细胞悬浮液(STE) 葡萄糖:提供等渗环境,增加溶液的粘度, 防止
DNA受机械作用而降解 Tris·HCl (三羟甲基氨甲烷) :
不含金属离子,与EDTA更能稳定DNA的活性 EDTA(乙二胺四乙酸二钠):
螯合Mg2+、Ca2+等金属离子,抑制DNase保 护DNA;EDTA的存在,有利于溶菌作用。
温放置5min;
(3)加入200l细菌裂解液,颠倒混匀,待溶液变粘稠后,
立即进行下一个步骤; (4)加入150l中和液,上下颠倒混合后置冰上 5min,
12,000转/分,离心 5min;
(5)将上清移至另一管中,加入等体积的TE饱和酚/氯仿
(1:1)混和液,振荡混匀1min,12,000转/分离心 2min;
离心 2min
+等体积TE
饱和酚/氯仿
移上清时要定量,不要混入白色沉淀物或漂浮物
饱和酚/氯仿(1:1)
!!带手套,一旦沾染皮肤请立即用大量清水冲洗! 酚:使蛋白质变性沉淀 变性效果好,但与水部分互溶。
氯仿:可使蛋白质变性沉淀 变性效果没有酚好,但与水不相混溶。
离心后分层
上层水相含质粒 水相下面是变性蛋白 下层酚/氯仿
冰上 5min后 12,000转/分
离心 5min
移上层水相
于另一管中
7
接提取质粒的操作流程及具体原理
(包括一些试剂的作用原理 )
移上层水相
于另一管中
分层
+等体积 氯仿/异戊醇
!!移上清时要定量,宁少勿多!
氯仿/异戊醇(24:1)溶液
氯仿:将残留在水相中的酚带走; 也可以再次使蛋白变性。
异戊醇:降低分子表面张力,减少气泡产生;
重组质粒DNA的提取及酶切鉴定

实验七重组质粒DNA的提取及酶切鉴定【实验原理】分离制备质粒DNA的方法很多,其中常用的方法有碱裂解法、煮沸法、SDS法、羟基磷灰石层析法等。
在实际操作中可以根据宿主菌株类型、质粒分子大小、碱基组成和结构等特点以及质粒DNA的用途进行选择。
本实验采用SDS碱裂解法提取重组质粒DNA,十二烷基磺酸钠(SDS)是一种阴离子表面活性剂,它既能使细菌细胞裂解,又能使一些蛋白质变性。
限制性内切酶能特异地结合于一段被称为限制性酶识别序列的DNA序列之内或其附近的特异位点上,并切割双链DNA。
限制性内切酶识别序列长度一般为4~8个呈回文序列的特异核苷酸对。
由于限制性内切酶的切割特性不同,分子生物学中主要用到Ⅱ型限制性内切酶(切割位置在识别序列内部)。
对质粒进行酶切,通过跑胶观察片段大小,从而鉴定质粒。
【实验步骤】本次实验所用的质粒提取试剂盒为天根的质粒小提试剂盒,操作步骤按说明书进行。
1. 吸附柱中加500ul 平衡液(BL),12000rpm离心1min ,弃收集管中的液体。
2.取1.5ml菌液至2ml离心管中,12,000rpm离心1min,弃上清。
3. 加250ul solution Ⅰ(P1),vortex。
4. 加250 solution Ⅱ(P2),上下颠倒混匀。
操作时间不能超过5min 注:此步骤不宜超过5 min。
5. 加350 solution Ⅲ(P3),立即颠倒混匀几次。
12000rpm离心10min。
6. 吸取上清加入吸附柱中,尽量不要吸出沉淀12000rpm离心1min ,弃收集管中的液体。
注:此时4℃离心不利于沉淀沉降。
7. 加入600μL漂洗液(PW)于离心吸附柱中,12000rpm离心1min ,倒掉废液。
8. 重复上一步,9. 空管离2min。
将吸附柱放入1.5ml离心管中,在超净台中晾5min。
10. 将700 μl的Rinse B加入Spin Column中,12,000 rpm离心30 sec,弃滤液。
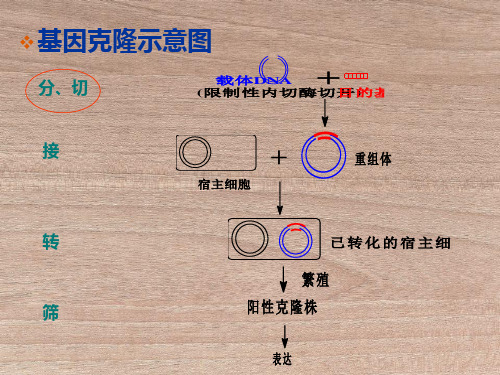
重组质粒的酶切和PCR验证

重组质粒的酶切和PCR验证⼭东⼤学实验报告 2013年 11 ⽉26⽇⾄12⽉10⽇姓名系年级 2011级⽣物技术组别四科⽬分⼦实验同组者学号题⽬重组质粒的酶切和PCR验证⼀、【实验题⽬】重组质粒的酶切和PCR验证⼆、【实验⽬的】1.学习和掌握PCR反应的基本原理和实验技术。
2.了解引物设计的基本要求。
3.复习凝胶电泳进⾏DNA分离纯化的⽅法。
4.复习DNA的酶切操作技术。
三、【实验试剂及器材】试剂:Hin d III 、酶切Buffer、Hin d Ⅲ、ddH2O、琼脂糖,TAE缓冲液、Loading Buffer、EB 染液、PCR buffer,dNTPs,Taq DNA聚合酶,引物器材:枪和枪尖、1.5 ml的Ep管、电泳仪、PCR反应⼩管四、【实验原理】1.PCR技术原理PCR (po lymerase chain react ion) 中⽂译为聚合酶链式反应或体外扩增技术, 是1985 年美国Cetus 公司⼈类遗传部、加利福尼亚⼤学洛杉机分校和How ghes 医院等联合建⽴的⼀项⽣物技术. 其原理类似于天然DNA 的复制, 在模板DNA、引物和4 种dN TP 等存在的条件下,依赖于DNA 聚合酶(Taq 酶) 的酶促合成反应. 其具体反应分三步: 变性、退⽕、聚合. 以上三步为⼀个循环, 如图1. 每⼀循环的产物DNA ⼜可以作为下⼀个循环的模板, 数⼩时后, 介于两个引物之间的⽬的DNA 得到⼤量的复制, 经过25~ 30 次循环, DNA 数量可达2×106-7拷贝数。
PCR基本原理是以单链DNA为模板,4种dNTP为底物,在模板3’末端有引物存在的情况下,⽤酶进⾏互补链的延伸,多次反复的循环能使微量的模板DNA得到极⼤程度的扩增。
在微量离⼼管中,加⼊与待扩增的DNA⽚段两端已知序列分别互补的两个引物、适量的缓冲液、微量的DNA膜板、四种dNTP溶液、耐热Taq DNA聚合酶、Mg2+等。
实验一二重组质粒DNA提取双酶切鉴定凝胶电泳紫外分光法
制胶的桌面相对水平。倒胶时尽量减少气泡的产 生。EB如果在制胶时加入,在60度左右时加入, 使终浓度为0.5ug/ml。不宜过低,染色成像不明显;
不宜过高,导致背景太深。摇匀要沿着瓶壁摇动, 尽量减少气泡产生的可能性。高浓度胶例如2%以 上的EB很难摇匀,而且凝的速度也相对快,强烈 建议跑完胶之后再用EB染色。
31
2.4 双酶切体系
合计
质粒DNA 10×M buffer BamHⅠ HindⅢ ddH2O
37 ℃,1-2h
10μl 2μl 1μl 1μl 6μl
20μl
同学自己加
准备室 老师已 加好
32
思考题?
❖ 为什么要对重组质粒DNA进行双酶切?
33
三、酶切质粒DNA后的琼脂糖凝胶 电泳
1 原理
上层水相转移到新EP管,加入2倍体积无水乙醇,颠倒混匀, -20 ℃ 冰箱中放置15min,12000g10min
弃上清,沉淀中加1ml 70%乙醇,12000g×5min 去上清,管平放室温静置10min, 20 µl TE 溶解DNA
RCF = 1.12r(rpm/1000)2
RCF: 离心力(g) r: 离心半径(mm) rpm: 每分钟转速(r/min)
4.室温凝胶30分钟
过程中不要碰到梳子,尽量保持胶的位置不移动。 时间不宜过久,导致胶干燥变形;不宜过短,影 响胶内部孔径形成。
5.拔梳子,放入电泳槽。
缓缓地将梳子垂直从梳孔拔出,尽可能使梳子是 同时从各个胶孔拔出的。暂时不用的胶最好放入 电泳槽电泳液中浸泡。电泳液要浸没胶1mm。
三、上样电泳
步骤
Buffer不要用成H2O,称量相对准确
2.融胶,加热到胶产生大量的气泡时,拿出摇匀, 非常热,小心烫手,另外注意不要加热过度使胶 继续加热到完全溶解,拿出摇匀,再加热到沸腾。 冲出瓶子。因此注意选择起码为胶体积2倍以上的
重组质粒地酶切鉴定和PCR实验

重组质粒的酶切鉴定与PCR实验一、【实验目的】1、酶切鉴定重组质粒插入片段的大小;2、学习和掌握PCR反响的根本原理和操作技术,了解引物设计的根本要求。
二、【实验原理】1、PCR反响根本原理PCR技术的根本原理类似于DNA的天然复制过程,其特异性依赖于与靶序列两端互补的寡核苷酸引物。
PCR由变性--退火--延伸三个根本反响步骤构成:①模板DNA的变性:模板DNA经加热至93℃左右一定时间后,使模板DNA双链或经PCR扩增形成的双链DNA解离,使之成为单链,以便它与引物结合,为下轮反响作准备;②模板DNA与引物的退火〔复性〕:模板DNA经加热变性成单链后,温度降至55℃左右,引物与模板DNA单链的互补序列配对结合;③引物的延伸:DNA模板--引物结合物在TaqDNA聚合酶的作用下,以dNTP为反响原料,靶序列为模板,按碱基互补配对与半保存复制原理,合成一条新的与模板DNA链互补的半保存复制链,重复循环变性--退火--延伸三过程就可获得更多的“半保存复制链〞,而且这种新链又可成为下次循环的模板。
每完成一个循环需2~4分钟,2~3小时就能将待扩目的基因扩增放大几百万倍。
PCR反响原理图2、PCR反响体系与反响条件(1) 标准的PCR反响体系①PCR反响的缓冲液提供适宜的酸碱度与某些离子③底物浓度 dNTP以等摩尔浓度配制,20~200umol/L④TaqDNA聚合酶 2.5U(100ul〕⑥反响温度和循环次数变性温度和时间 95℃,30s退火温度和时间低于引物Tm值5 ℃左右,一般在45~55℃延伸温度和时间 72℃,1min/kb(10kb内〕Tm值=4(G+C) +2(A+T)循环次数:一般为25 ~ 30次。
循环数决定PCR扩增的产量。
模板初始浓度低,可增加循环数以便达到有效的扩增量。
但循环数并不是可以无限增加的。
一般循环数为30个左右,循环数超过30个以后,DNA聚合酶活性逐渐达到饱和,产物的量不再随循环数的增加而增加,出现了所谓的“平台期〞。
重组质粒鉴定

重组质粒进行鉴定时,可以采用两种方法进行鉴定。
1.通过pcr方法鉴定:以重组质粒为模板,pcr产物的特异性引物或载体的通用引物进行PCR 扩增后电泳鉴定。
2..就是酶切鉴定:双酶切鉴定时只要出现质粒条带和你的插入片段的目的条带就行了。
至于出现质粒条带很亮,而目的条带暗的现象,其实很正常。
因为一般情况下,质粒的碱基数比你的目的条带的碱基数多的多(一般质粒碱基都有好几千bp,而目的条带通常就几百到一千多bp)。
当我们用EB进行染色时,EB是掺入到到dna链中,碱基数越多则掺入的eb就越多,在紫外光下显示的条带就越亮,也就是说条带亮度与你的片段的长度成正比。
最后,如果两种方法都鉴定正确了,你就可以送到公司进行测序,做最后的鉴定了。
如果你非要看到你的目的条带很明显的话,也可以采取如下方法:
1.电泳时吸取的产物量加大,加入到大孔梳子的胶当中,如可以加产物10微升或更多。
2.凝胶成像拍照时,可以适当把曝光时间提高一点。
3.如果还是不清楚,就把你的酶切产物浓缩一下。
质粒重组实验报告

质粒重组实验报告质粒重组实验报告引言:质粒重组是一种重要的实验技术,可以将外源基因导入到宿主细胞中,用于研究基因功能、蛋白质表达等方面。
本实验旨在通过质粒重组技术,将外源基因成功导入到大肠杆菌中,并通过酶切、连接、转化等步骤验证重组质粒的构建。
材料与方法:1. 外源基因:选择了编码了绿色荧光蛋白(GFP)的质粒pGFP。
2. 宿主细胞:采用大肠杆菌作为宿主细胞。
3. 酶切酶:使用限制酶EcoRI和BamHI进行酶切反应。
4. 连接酶:使用T4 DNA连接酶进行连接反应。
5. 培养基:含有抗生素的LB培养基。
实验步骤:1. 酶切反应:将pGFP质粒和EcoRI、BamHI酶切酶按照指定条件进行酶切反应,以得到线性化的质粒和目标基因片段。
2. 连接反应:将线性化的质粒和目标基因片段加入连接酶缓冲液中,按照指定条件进行连接反应,形成重组质粒。
3. 转化:将重组质粒加入含有大肠杆菌的培养基中,通过热激转化法使质粒进入细胞。
4. 筛选:将转化后的细胞均匀涂布在含有抗生素的LB平板上,筛选出含有重组质粒的菌落。
5. 验证:通过PCR、酶切等方法对筛选出的菌落进行验证,确认质粒重组成功。
结果与讨论:经过酶切、连接、转化等步骤,成功构建了质粒重组实验系统。
在LB平板上筛选出了多个抗生素耐受的菌落,表明转化成功。
通过PCR验证,检测到了目标基因片段的特异扩增带,证明重组质粒中成功导入了外源基因。
进一步进行酶切验证,发现重组质粒经过连接反应后,酶切位点发生改变,与未连接的质粒有明显差异。
这些结果表明,质粒重组实验成功构建了重组质粒,并将外源基因导入到大肠杆菌中。
质粒重组实验的成功对于基因功能研究和蛋白质表达具有重要意义。
通过重组质粒,可以将外源基因导入到宿主细胞中,进而研究其在生物体内的功能和表达情况。
例如,可以通过重组质粒实现基因敲除、基因表达调控等功能,从而深入了解基因在生物体内的作用机制。
同时,重组质粒还可以用于蛋白质表达,通过外源基因的转录和翻译,使宿主细胞产生目标蛋白质,用于研究其结构、功能等方面。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
上样图示
上样和电泳: 每组上样2个( 未酶解质粒
和双酶切产物) 未酶解质粒10 μL +2 μL凝胶加样缓冲液 双酶切产物20 μL +4 μL凝胶加样缓冲液 Marker 5 μL, 放中间,2组1块胶 各组做好准备,两组统一上样后电泳 电泳槽和电泳仪不可移动 10 μL 20 μL +2 μL +4 μL 5 μL 10 μL 20 μL +2 μL +4 μL
2. 质粒DNA的限制性内切酶的双酶切
注意: 1. 注意加样顺序 2. 保证每样试剂加入反应体系 3. 加样完成后需混匀,短暂离心 4. 反应结束后,上样电泳前需短 暂离心,再加凝胶加样缓冲液 5. 及时将酶放回-20 ℃冰箱以保 证酶的活力
Buffer 4
BamHⅠ(20U/ μl) XbaⅠ(20 U/ μl) dd H2O 质粒DNA
将细菌培养物全部倒入15mL离心管,4500rpm,5min 请保持同步! 弃上清,沥干,注意细菌沉淀别倒掉了! 离心管配平 加250 μl Buffer S1,吹打混匀(也可Vortex)以充分悬浮细菌,转入1.5ml离心管 加250 μl Buffer S2,上下颠倒,温和混匀4~6次使菌体裂解 避免剧烈摇晃,此步骤不宜超过5min 加350 μl Buffer S3,上下颠倒,温和混匀6~8次,12000g 离心10min 避免剧烈摇晃 吸取上清至已置于 2ml 离心管的制备管中,12000g离心1min,弃滤液 同上,在制备管中加500 μl Buffer W1, 12000g离心1min ,弃滤液 在制备管中加700 μl Buffer W2, 12000g 离心1min ,弃滤液, 重复以Buffer W2洗涤(离心)一次,再空离心1次 Eluent液65 ℃温育可提高洗脱效率 将制备管移入新的1.5ml离心管,在膜中央加60 μl Eluent或去 离子水,室温静置1min, 12000g离心1min,即为质粒DNA。
凝固后成凝胶
标准琼脂糖 低熔点琼脂糖
DNA在凝胶中的迁移率的影响因素
DNA分子的大小 琼脂糖的浓度
DNA的构象
所加电压 嵌入染料的存在 电泳缓冲液的组成及其 离子强度
DNA条带迁移距离(cm) ( 一般采用相对迁移率会更好)
不同含量标准的琼脂糖凝胶的分离范围
琼脂糖凝胶浓度 (%) 0.3 0.5 0.6 0.7 1.0 1.2 1.5 2.0 DNA分子分离范围 (kb) 5~60 1~30 1~20 0.8~12 0.5~10 0.4~6 0.2~4 0.1~3
加入TAE 60ml,勿晃
1. 标液面高度以观察水分是否蒸发过多,若
溶解后水分蒸发过多,补水
2. 三角烧瓶上覆打过洞的保鲜膜(防水蒸发) 3. 防爆沸和烫伤(带棉手套) 4. 加6μLGelRed后立即灌胶(戴手套) 微波炉溶胶 注意先做好制胶准备!
在电泳槽内加电泳缓冲液 拔去加样梳,将制好的凝胶连同内架 放入电泳槽内,保证液面高于凝胶面 加入6 ×凝胶加样缓冲液和 DNA(1:5比例混合)
实验二 重组DNA的提取 及酶切鉴定
1
实验目的
掌握以下方法和技术:
1. 质粒DNA的小量快速制备
2. 质粒DNA的限制性内切酶酶切
3. DNA的琼脂糖凝胶电泳
2
实验原理
1. 质粒DNA的小量快速制备
分离质粒DNA有三个步骤:
1. 培养细菌使质粒扩增
2. 收集和裂解细菌(本实验:SDS裂解)
DNA分子中的磷酸二酯键骨架在盐溶液 的作用下脱水,使得暴露的磷酸基团吸附硅
胶。双链DNA分子一旦被硅胶吸附,一般的
洗液不能将其洗脱下来,只有水溶性的缓冲 液,如TE或水重新水化后,DNA才能从层 析柱上定量回收。
2. 质粒DNA的限制性内切酶酶切
c-myc基因片段4.8 kb
BamH I + Xba I双酶切(酶切完 全时) 琼脂糖凝胶电泳凝胶中2条区带 3.5kb
感谢您的聆听
DNA从阴极向阳极迁移
依次加样,并加好Marker 连通电源,80~100V电泳至 溴酚蓝带达到凝胶的1/2处 结果观察和记录
GelRed从阳极向阴极迁移
(加样后不能移动电泳槽!)源自凝胶成像仪3. DNA的1%琼脂糖凝胶电泳
制胶:1% agarose ,防过热暴沸 上样准备 (1份凝胶加样缓冲液加5份 DNA) 未酶切 酶切 Marker 未酶切 酶切
1. 首次使用前,将RNaseA全部加入Buffer S1中,4 ℃贮存
2.首次使用前,Buffer W2 concentrate 中加入指定体积的无水乙醇
3. 使用前检查Buffer S2是否出现沉淀,若有应在37℃加热溶解并冷却 至室温使用 4. 自行准备无核酸和核酸酶污染的tip头和离心管 5. Buffer S2使用后立即盖紧瓶盖,以免空气中的CO2中和Buffer S2中 的NaOH,降低溶菌效率
Xba I
pSV-c-myc
BamH I
pSV2载体片段 c-myc 目的基因片段
4.8kb
pSV2载体片段3.5kb
3. 琼脂糖凝胶电泳
琼脂糖-海藻中提 取的线状高聚物
不同类型琼脂糖的特性
琼脂糖类型 凝结温度/℃ 熔化温度/℃
35~38 25~35 90~95 63~65
加热融化成清澈、 透明的溶液
2 μL
1 μL 1 μL 10 μL 6 μL 20 μL
混匀后短暂离心,37 ℃水浴1小时
准备好制胶器,插好梳子并保证 梳子距底部0.5mm-1.2mm
准备好凝胶,加热熔解
灌胶(事先加GelRed)注意赶气泡
待凝
1%agar ose凝 胶
1%agar ose凝 胶
称量agarose
注意事项:
DNA琼脂糖凝胶电泳缓冲液
缓冲液 类型 Ⅰ
成分 0.25%溴酚蓝 0.25%二甲苯氰FF 40%蔗糖 0.25%溴酚蓝 0.25%二甲苯氰FF 15%Ficoll 0.25%溴酚蓝 0.25%二甲苯氰FF 30%甘油 0.25%溴酚蓝 40%蔗糖
贮存温度 4℃
Ⅱ
室温
Ⅲ
4℃
Ⅳ
4℃
质粒DNA的3种构象
3. 分离和纯化质粒DNA(本实验:碱变性法)
碱变性法抽提质粒DNA
Solution II Solution III 离心后去向 pH12.6 pH4.8
染色体 DNA
氢键断裂, 双螺旋解开
不能复性
沉淀中
氢键部分断 裂,cccDNA 质粒DNA 互补链不完 全分离
复性
上清中
硅基质材料(树脂)与DNA双链相互作用的原理
-23130 -9416 -6557 -4361 -2322 -2027
1%agarose凝胶电泳
发散性思考: 1. 为何细菌平板上的长出的细菌集合是单克隆 (一个细菌来源)的?请例举可能的解释来支 持或反对这一说法。
2. 为何线性的质粒不易转入细菌内,如何从机理 上解释,从实验中证明?连接时是否会出现多 聚体?
4
实验结果观察和记录
80~100V电泳,直到溴酚蓝带泳动到凝胶的1/2至2/3处停止。 紫外灯下观察,拍照记录。
预期结果
实验组 对照组未酶切 未酶切 对照组 实验组 质粒 双酶切 双酶切Marker双酶切 双酶切 质粒
未酶切 对照组 实验组 λDNA/Hind ш 质粒 双酶切 双酶切 Marker
未酶切 λDNA/Hind ш 质粒 Marker
-23130 -9416 -6557 -4361 -2322 -2027
琼脂糖凝胶电泳凝胶中出现1或2或 3条带都有可能
1%agarose凝胶电泳
存在复制中间体可能
超螺旋的质粒跑得比线性快还是慢?
3
实验步骤
1. 质粒DNA的小量快速制备
(AXYGEN AxyPrep质粒DNA小量制备试剂盒)注意事项: