真核生物之酵母表达系统
酵母细胞的基因表达调控和代谢调节研究

酵母细胞的基因表达调控和代谢调节研究酵母细胞作为微生物中的重要代表,是一类单细胞真核生物,其广泛应用于生物学、发酵工艺和医学研究中。
酵母细胞的基因表达调控和代谢调节是酵母细胞研究的重要方向,也是生物学研究的重要领域,对理解生命过程以及人类疾病的发病机制有着重要的意义。
一、基因表达调控的研究酵母细胞具有很强的遗传可控性,基因表达调控的研究一直是酵母细胞研究的热点之一。
酵母细胞中基因表达调控主要包括转录前、转录后和调节因子等方面的研究。
1、转录前调控转录前调控主要是指通过转录因子对基因启动子的结合来调控基因的转录速率。
酵母细胞的转录因子主要包括激活转录因子和抑制转录因子两类。
激活转录因子通过结合启动子使得转录因子蛋白与RNA聚合酶共同形成转录复合物,从而促进基因的转录;抑制转录因子则通过结合启动子抑制转录复合物的形成,使得基因的转录过程被抑制。
2、转录后调控转录后调控主要是指转录后修饰以及mRNA剪接等方式对RNA的降解速率和转录的抑制作用。
酵母细胞中常见的转录后修饰有剪切、RNA编辑、加工等方式,这些修饰可以影响RNA的稳定性、翻译速率和结构功能等方面。
3、调节因子调节因子是指在DNA核苷酸序列中通过某种方式调节基因表达的核酸分子。
调节因子可以增强或者抑制某个基因的转录,也可以同时调控多个基因的转录。
酵母细胞中的调节因子主要有染色质重塑复合体、DNA甲基转移酶等。
二、代谢调节的研究代谢调节是指在代谢途径中通过调节酵素活性和代谢产物浓度等方式影响代谢反应的调节过程。
酵母细胞的代谢途径包括葡萄糖酵解、异养代谢、脂质代谢、氨基酸代谢等。
1、葡萄糖酵解葡萄糖酵解是指将葡萄糖分解为乳酸或乙醇代谢的过程。
酵母细胞的葡萄糖酵解主要受到两种调控方式的影响,一种是通过启动子上的转录因子对基因表达的调节,另一种是通过一些代谢产物对基因表达的反馈调控。
2、异养代谢异养代谢是指在缺少氧气或碳源时,酵母细胞通过吸收硫酸盐、氨、磷酸盐等原始无机物质方式进行代谢的过程。
酵母表达系统

4、甲醇酵母系统高效表达影响因素与对策
载体稳定性 基因剂量 整合位点 甲醇利用表型 mRNA5’端 AT含量分泌信号 表达产物稳定性
1)载体稳定性
同拷贝数时,整合型的比自主复制型的表达水平高 YRp型载体的稳定化:
选择—非选择培养交替数十代可得稳定的整合子 ,但费时,整合位点不确定。 采用YIp型载体: 更易实现整合、整合位点清楚
2)基因剂量
外源基因表达存在基因剂量效应 筛选多拷贝整合子
载体引入G418/Zeocin抗性标记,整合子拷贝数 与抗性成正相关,采用高G418/Zeocin抗性转化子。 体外串联多个表达盒,直接获多拷贝整合子 采用YRp型载体稳定化技术获高拷贝整合子 构建高拷贝整合型表达载体
3)整合位点
外源基因表达盒整合于AOX/MOX或标记基因处,均 可高效表达 毕赤酵母中个别情况整合于His4位点的比AOX1位点 的低
2)分泌表达产物过糖基化
(二) 甲醇酵母表达系统
甲醇酵母与甲醇氧化酶启动子 甲醇酵母表达系统的优缺点 甲醇酵母表达系统操作原理 甲醇酵母系统高效表达影响因素与对策 甲醇酵母表达系统的应用
1、甲醇酵母与甲醇氧化酶启动子
甲醇酵母(methylotrophic yeast) 指可利用甲醇作单一碳源的一类酵母。 毕赤酵母(Pichia pastoris) 汉森酵母(Hansenula ploymorpha) 假丝酵母(Candia boidinii)
组成的、复杂分支结构的现象。增加了免疫原性、对活 性与药代稳定性均有影响。 *糖链组成
O型糖链仅由甘露糖组成、而哺乳细胞的还含唾液酸 基团
4、酿酒酵母表达系统的缺陷
1)表达水平普遍不高 A、表达载体传代不稳定(YEp、YRp) B、所采用的强启动子调控不严谨 C、不能利用简单的无机培养基进行高密度发酵
蛋白质表达系统介绍不同的蛋白质表达系统及其优缺点

蛋白质表达系统介绍不同的蛋白质表达系统及其优缺点蛋白质表达是生物学研究中一项重要的技术,它可以通过合成蛋白质来研究其结构和功能。
蛋白质表达系统是实现这一过程的关键工具,主要包括原核表达系统和真核表达系统两种。
本文将对这两种蛋白质表达系统进行介绍,并分析它们的优缺点。
一、原核表达系统原核表达系统是利用原核生物(如大肠杆菌)来表达外源蛋白质的系统。
该系统具有以下特点:1. 高表达水平:大肠杆菌是常用的原核表达宿主,具有高表达水平的优势。
通过利用原核细胞的强大蛋白质合成机器,可以获得高产量的外源蛋白质。
2. 易操作性:原核表达系统相对简单,操作步骤少,易于操作和控制。
不需要复杂的细胞培养条件,可以在常见培养基中进行表达。
3. 快速表达:从启动表达到获得蛋白质通常只需要数小时至数天,速度较快。
这使得原核表达系统在高通量表达和快速实验中具有优势。
然而,原核表达系统也存在一些缺点:1. 外源蛋白质折叠问题:由于原核细胞的机器无法正确折叠某些复杂蛋白质,这可能导致外源蛋白质的不正确折叠和失活。
2. 原核特异性翻译后修饰:原核细胞缺乏一些真核细胞所具有的翻译后修饰机制,这可能影响蛋白质的功能和稳定性。
3. 复杂蛋白质表达困难:对于复杂蛋白质(如膜蛋白),原核表达系统通常无法达到理想的表达水平和正确的折叠结构。
二、真核表达系统真核表达系统主要利用真核生物(如酵母、昆虫细胞和哺乳动物细胞)来表达外源蛋白质。
真核表达系统具有以下特点:1. 正确的折叠和修饰:真核细胞具有复杂的蛋白质折叠和修饰机制,能够产生正确折叠和修饰的蛋白质。
2. 适用于复杂蛋白质:真核表达系统适用于复杂蛋白质(如膜蛋白)的表达。
真核细胞提供了正确的环境和细胞器,能够较好地表达这类蛋白质。
3. 适用于大规模表达:真核细胞通常可以进行大规模培养和表达,适用于工业化生产。
然而,真核表达系统也存在一些缺点:1. 低表达水平:相对于原核表达系统,真核表达系统的表达水平较低,可能无法满足高产量蛋白质的需求。
酵母表达(1)

酵母表达引言酵母是一类单细胞真核生物,被广泛应用于生物学研究中。
酵母表达系统是指利用酵母细胞表达外源基因的技术,被广泛应用于蛋白质的高效表达和产量大规模生产。
本文将介绍酵母表达系统的原理、优势和应用。
原理酵母表达系统的核心原理是将外源基因导入酵母细胞,并通过酵母细胞的转录、翻译和修饰机制,使外源基因在酵母细胞中得到表达和功能发挥。
通常情况下,酵母表达系统主要采用酵母菌属的酿酒酵母(Saccharomyces cerevisiae)或毕赤酵母(Pichia pastoris)作为宿主细胞。
1.酵母转录机制:酵母细胞的基因表达主要通过RNA聚合酶Ⅱ进行转录,产生mRNA分子。
2.酵母翻译机制:酵母细胞通过核糖体进行翻译,将mRNA翻译成蛋白质。
3.酵母修饰机制:酵母细胞具有多种修饰酶,可以对蛋白质进行翻译后修饰,如糖基化、磷酸化等。
优势相比其他常用的表达系统,酵母表达系统具有一系列的优势:1.高效表达能力:酵母表达系统能够实现高水平的外源基因表达,产量可达到克级。
2.翻译后修饰:酵母细胞具有多种修饰酶,可以对蛋白质进行翻译后修饰,使蛋白质得到正确的糖基化等修饰。
3.生长条件简单:酵母菌生长条件相对简单,可以在常规培养基中进行培养,对培养条件的要求相对较低。
4.可溶性蛋白质表达:酵母细胞具有较强的蛋白质折叠和修饰能力,能够高效地表达可溶性蛋白质。
应用酵母表达系统广泛应用于以下领域:1.蛋白质研究:酵母表达系统可用于大规模蛋白质表达和纯化,为蛋白质的结构、功能和相互作用研究提供了高效的工具。
2.药物筛选:酵母表达系统可用于药物靶点鉴定和药物分子筛选,加速药物研发过程。
3.疫苗研究:酵母表达系统可用于疫苗候选抗原的高效表达和产量大规模生产。
4.代谢工程:酵母表达系统可用于代谢工程领域,利用酵母细胞对外源代谢产物的高效合成能力,实现产生复杂化合物的目标。
5.生物制药:酵母表达系统已经被广泛应用于生物制药领域,用于生产重组蛋白和抗体等生物药物。
酵母表面展示技术.pptx
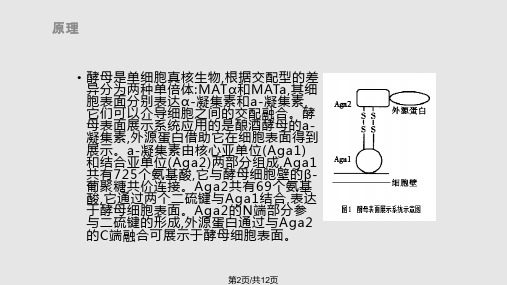
• 载体蛋白
酵母细胞表面包被着一层坚硬的细胞壁,由内 层的葡聚糖骨架和外层的甘露糖蛋白构成。甘 露糖蛋白主要包括两类蛋白质:一种以非共价 方式松散地结合于细胞壁,可以用SDS 抽提; 另一类蛋白必需以 β-1,3 或β-1,6 葡聚糖酶消 化细胞壁后才能抽提,这类蛋白常含有 GPI 锚 定区域。α-凝集素和絮凝素以及细胞壁蛋白如 Cwp1p,Cwp2p,Tip1p 等都属于 GPI 家族 蛋白,外源蛋白与它们融合后可被共价锚定于 细胞表面,这些蛋白是常用的酵母表面展示载 体蛋白。
第6页与外源蛋白融合,并将融合蛋白展露表达 在细胞表面,此系统又包括两个子系统: GPI锚定系统和絮凝结构域锚定系统。 GPI锚定系统是利用絮凝素Flo1p的C末端含有的GPI信号锚定外源蛋白,此系 统与凝集素展示相似; 絮凝结构域锚定系统是利用Flo1p的中间絮凝功能结构 域与外源蛋白融合,通过絮凝功能结构域识别酵母细胞壁中的甘露聚糖链并以 非共价作用诱导细胞粘附、聚集成可逆性絮状物
第10页/共12页
• 酵母表面展示与酶技术 • 酶的固定化是指借助物理或者化学方法将酶固定于特殊
的相,使得酶与整体流体分开,但是仍然能够进行底物 和效应物分子交换并发挥其催化效能的一种技术。与游 离酶相比,固定化酶提高了酶的稳定性,并使酶能够反 复回收利用。但是,传统的固定化方法也会产生一些不 利因素,例如由于增加固定化操作,导致酶固定化过程 中的活性收率损失;另外由于固定化操作需用载体,因 而增加了载体成本费和固定化操作费用。利用表面展示 技术将具有催化活性的酶展示于酵母等微生物细胞表面 就形成了全细胞催化剂,与传统的细胞内酶和外分泌酶 不同,表面展示的酶以共价或非共价方式固定于细胞外 表面,这种独特的空间定位使其相对自由酶而言有许多 优良的特性,如温度、有机溶剂稳定性、可多次重复使 用等,这些特点与传统的固定化酶技术相似,但无需额 外的蛋白纯化和固定的操作,有着良好的应用前景。
酵母表达系统使用心得

Pichia酵母表达系统使用心得甲醇酵母表达系统有不少优点,其中以Invitrogen公司的Pichia酵母表达系统最为人熟知,并广泛应用于外源蛋白的表达。
虽然说酵母表达操作简单表达量高,但是在实际操作中,并不是每个外源基因都能顺利得到高表达的。
不少人在操作中会遇到这样那样的问题,收集了部分用户在使用EasySelect Pichia Expression System这个被誉为最简单的毕赤酵母表达的经典试剂盒过程中的心得体会。
其中Xiang Yang是来自美国乔治城大学(Georgetown University)Lombardi癌症中心(Lombardi Cancer Center),部分用户来自国内。
甲醇酵母部分优点:1.属于真核表达系统,具有一定的蛋白质翻译后加工,有利于真核蛋白的表达;2.AOX强效启动子,外源基因产物表达量高,表达产物可以达到每升数克的水平;3.酵母培养、转化、高密度发酵等操作接近原核生物,远较真核系统简单,非常适合大规模工业化生产;4.可以诱导表达,也可以分泌表达,便于产物纯化;5.可以甲醇代替IPTG作为诱导物,部分甲醇酵母更可以用工业甲醇替代葡萄糖作为碳源,生产成本低。
产品性能:优点——使用简单,表达量高,His-tag便于纯化;缺点——酵母表达蛋白有时会出现蛋白切割问题。
巴斯德毕赤酵母(Pichia pastoris)是一种能高效表达重组蛋白的酵母品种,一方面由于其是属于真核生物,因此表达出来的蛋白可以进行糖基化修饰,另一方面毕赤酵母生长速度快,可以将表达的蛋白分泌到培养基中,方便蛋白纯化。
毕赤酵母表达载体pPICZ在多克隆位点(MCR)3'端带有his-tag和c-myc epitopes,这些tag有利于常规检测和纯化,而且在MCR5'端引入了alpha factor(α-factor)用以分泌表达,并且在表达后α-factor可以自动被切除。
第十五章:酵母菌基因工程选编

③易进行载体DNA的导入。DNA转化技 术的不断发展优化,多数酵母菌可 以取得较高的转化率;
④培养条件简单,容易进行高密度 发酵;
⑤能将外源基因表达产物分泌到培 养基中;
⑥有类似高等真核生物的蛋白质翻 译后的修饰功能。
2.缺陷在于:
①表达效率相对低; ②酵母常有密码子偏爱性,真核基
因在其中表达时需要人工修正。
2.含有ARS的YRp和YEp质粒及其构建
①ARS为酵母菌中的自主复制序列,大 小在0.8-1.5Kb,染色体上每30-40bp 就有一个ARS元件。
②由染色体ARS构成的质粒称为YRp,而 由2μ质粒构建的杂合质粒为YEp。
③上述两类质粒在酿酒酵母中的拷贝数 最高可达200个,但是经过几代培养 后,质粒丢失率达50%-70%,主要由 于分配不均匀所致。
三.抑制超糖基化作用的突变宿主菌
许多真核生物的蛋白质在其天门冬 酰胺侧链上接有寡糖基团,常常影 响蛋白质的生物活性。整个糖单位 由糖基核心和外侧糖链两部分组成。
酵母菌普遍拥有完整的糖基化系统,酿 酒酵母细胞内的天门冬酰胺侧链糖基修 饰和加工系统对来自高等动物和人的异 源蛋白活性表达是极为有利的,但野生 型酿酒酵母对异源蛋白的糖基化反应很 难控制,呈超糖基化倾向,因此超糖基 化缺陷菌株非常重要。
②YAC载体的装载量建
①YIP 载体由大肠杆菌质粒和酵母的 DNA 片段组成,可与受体或宿主的染色体 DNA 同源重组,整合进入宿主染色体中,酵母 片段只提供选择性标志,没有复制起点。
②转化率低(只有1-10转化子/微克DNA), 但转化子遗传性稳定,多用于遗传分析。
一.广泛用于外源基因表达的酵母宿主菌
目前已广泛用于外源基因表达的研究的酵母菌包括:
基因工程:第四章-酵母基因工程

UBC4-UBC5双突变型:
UBC4-UBC5双突变型能大幅度削弱泛
素介导的蛋白降解。
7个泛素连接酶基因的突变对衰减蛋白 降解作用同样有效。
6、内源性蛋白酶缺陷型的突变宿主菌
酿酒酵母具有20多种蛋白酶 空泡蛋白酶基因PEP4野生型和
pep4-3突变株
B-半乳糖苷酶活性明显升高
(三) 酵母菌的载体系统
酵母基因工程
酵母菌作为外源基因表达受体菌的特征 酵母菌的宿主系统 酵母菌的载体系统 酵母菌的转化系统 酵母菌的表达系统 利用重组酵母生产乙肝疫苗
1974 Clarck-Walker和Miklos发现在多数酿酒酵母 中存在质粒。
1978 Hinnen将来自一株酿酒酵母的leu2基因导入 另一株酿酒酵母,弥补了后者leu2的缺陷, 标志着酵母表达系统建立。
酵母菌有4个泛素编码基因:
UBI1 编码泛素-羧基延伸蛋白52 对数生长期表达 稳定期关闭
UBI2 编码泛素-羧基延伸蛋白52 对数生长期表达 稳定期关闭
UBI3 编码泛素-羧基延伸蛋白76 对数生长期表达 稳定期关闭
UBI4 编码泛素五聚体
对数生长期关闭 稳定期表达
酵母菌有7个泛素连接酶基因:
UBC1、UBC2、UBC3、UBC4、UBC5、UBC6、UBC7
酵母菌表达外源基因的优势: 全基因组测序,基因表达调控机理清楚,遗传 操作简便。 具有真核生物蛋白翻译后加工修饰系统。 能将外源基因表达产物分泌至培养基中。 大规模发酵工艺简单、成本低廉。
不含特异性病毒、不产毒素,被美国FDA认定为 安全的基因工程受体系统。
酵母菌表达外源基因的缺点:
表达产物的糖基化位点和结构特点 与高等真核生物有差距。
特点:
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
3、甲醇酵母表达系统操作原理
宿主株与标记基因 甲醇酵母系统的整合事件 胞内表达与分泌表达
甲醇酵母系统宿主
二大宿主系统主要特点
项目 最适温度 最适pH值 甘油阻遏 毕赤酵母 30℃ 4.5 是 汉森酵母 37℃ 4.5 否
糖基化
高密度发酵
部分过度
100g/L
较正常
100g/L
甲醇酵母系统宿主
二大宿主系统主要选择标记
半乳糖诱导、葡萄糖抑制
GAL10 Promoter
GAL80
GAL4
UAS
GAL1
GAL7
GAL10
A、 将GAL4的启动子换成GAL10的诱导型强启动子 B、半乳糖诱导GAL4高表达,不受GAL80产物抑制,激活GAL1等高效转录
3)pho4TS-PHO5启动子
低温诱导、磷酸盐抑制 A、PHO5启动子在培养基缺磷酸盐时启动转录
宿主株:GS115、KM71
可插入位点: 5’AOX1 3’AOX1
TT
转化子: GS115:His+Mut+
KM71:His+Muts
4) 基因取代(GS115,AOX1+)
转化子:His4+Muts
汉森酵母系统的高拷贝整合型表达载体
高拷贝整合元件: A、高度重复序列:rDNA
提供多个整合位点
3、酿酒酵母糖基化系统
糖基化位点:Asn-X-Thr/Ser(X代表任何氨基酸)
N-型糖基化:天门冬酰氨酸残基上的酰胺氮进行糖
基化,对蛋白质的折叠、稳定性及活性较重要。
O-型糖基化:苏氨酸/丝氨酸上的羟基氧进行糖基化
酿酒酵母糖基化特点
* 过糖基化(超糖基化修饰):
N型或O型糖基的外链进一步形成庞大的、由甘露糖
甲醇酵母系统的整合事件
YRp型载体:汉森系统
传代不稳定,传代过程同源或非同源重组,高选择 压力迫使高拷贝数整合,可达100拷贝。
YIp型载体:
A、在靶序列处线性化载体DNA,诱导同源重组
B、有“插入”和“取代”二类整合模式
C、主要为单拷贝整合,1-10%为多拷贝整合
毕赤酵母系统模式表达载体
1)AOX1位点插入
AOX1与AOX2 *毕赤酵母和假丝酵母基因组存在二个AOX基因 AOX1、AOX2 *AOX1与AOX2基因97%同源 *AOX1 占主导地位,负责AOX 99%以上活性
1、甲醇酵母与甲醇氧化酶启动子
甲醇氧化酶启动子 A、目前已发现的、最强的真核启动子 B、严谨调控型启动子 AOX1:葡萄糖和甘油脱阻遏、甲醇诱导 MOX:葡萄糖阻遏、甘油脱阻遏、甲醇诱导
信号肽:MFα 标记:Kan
4、甲醇酵母系统高效表达影响因素与对策
载体稳定性 基因剂量
整合位点
甲醇利用表型 mRNA5’端 AT含量分泌信号 表达产物稳定性
1)载体稳定性
同拷贝数时,整合型的比自主复制型的表达水平高 YRp型载体的稳定化: 选择—非选择培养交替数十代可得稳定的整合子, 但费时,整合位点不确定。 采用YIp型载体: 更易实现整合、整合位点清楚
7)分泌信号
分泌表达效率与所用分泌信号高度相关
可采用酿酒酵母来源的MFα、PHO或SUC等的信号 肽,或野生型信号肽,但适用性要比较分析。
8)表达产物稳定性
分泌表达时,胞外蛋白酶是要影响因素
降低培养基pH值:蛋白酶在酸性条件下活性较低
培养基中添加蛋白水解产物:竞争性抑制
采用蛋白酶缺陷宿主株:如P.pastoris SMD1168
宿主 毕赤酵母 汉森酵母 选择标记 his4、G418、 Zeocin、Cu++ leu2、 his3 、ura3、 G418
甲醇酵母系统宿主
毕赤酵母系统主要宿主株
宿主株 Y11430 GS115 特点 野生型、Mut+ his4- 、Mut+
KM71 his4-、 aox1::ARG4 (AOX1-)、Muts SMD1168 his4- 、胞外蛋白酶缺陷,高分泌 MC1003 his4-、AOX1-、AOX2-,甲醇不生长
UAS
URS
TATA
2、TATA盒:富含AT;
3、UAS:上游激活序列;
5、DAS:下游激活序列
酿酒酵母表达系统常用启动子
1)糖酵解途径中关键酶的强启动子,受葡萄糖诱导:
甘油醛-3-磷酸脱氢酶基因GAPDH
磷酸甘油激酶基因PKG
乙醇脱氢酶基因ADH
2)半乳糖激酶启动子(GAL1)
半乳糖诱导、葡萄糖抑制
毕赤酵母中个别情况整合于His4位点的比AOX1位点 的低
4)甲醇利用表型
在分泌表达情况下,甲醇慢生长型更利于表达
5)MRNA5’端
mRNA5’端非翻译区(UTR)的组成与长度 应尽量与AOX1/MOX的一致
应尽量避免在UTR区出现AUG和次级结构
6)基因序列的AT含量 AT含量越高、越容易出现类似于终止子的结构
酵母表达系统
酿酒酵母表达系统 甲醇酵母表达系统 其它酵母表达系统
(一) 酿酒酵母表达系统
酿酒酵母系统启动子 酿酒酵母分泌系统 酿酒酵母糖基化系统 酿酒酵母表达系统存在的问题
1、酿酒酵母启动子
起始 位点 mRNA 40-120bp 20-40bp 100-1400bp 1、转录起始位点; 4、URS:上游阻遏序列 DAS 编码序列
1、甲醇酵母与甲醇氧化酶启动子
甲醇氧化酶 A、甲醇代谢关键酶,占可溶蛋白的30%以上 B、名称和拷贝数不同 毕赤酵母/假丝酵母 汉森酵母 醇氧化酶 甲醇氧化酶 alcohol oxidase,AOX methanol oxidase,MOX AOX1、AOX2 MOX
1、甲醇酵母与甲醇氧化酶启动子
组成的、复杂分支结构的现象。增加了免疫原性、对活
性与药代稳定性均有影响。
*糖链组成
O型糖链仅由甘露糖组成、而哺乳细胞的还含唾液酸
基团
4、酿酒酵母表达系统的缺陷
1)表达水平普遍不高
A、表达载体传代不稳定(YEp、YRp)
B、所采用的强启动子调控不严谨
C、不能利用简单的无机培养基进行高密度发酵 2)分泌表达产物过糖基化
酿酒酵母信号肽特点
*保守性低,大多异源宿主系统的信号肽不能互用
*信号肽结构:
信号肽剪切位点 Met
正电荷区
疏水区
极性区
目的蛋白
MF-α信号肽
*分泌效率高
*在酵母系统具有通用性
*88个残基组成
Met KEX2 DAP DAP来自-Lys-Arg-Glu-Ala-Glu-Ala目的蛋白 DAP:STE13编码的二肽酶
(二) 甲醇酵母表达系统
甲醇酵母与甲醇氧化酶启动子 甲醇酵母表达系统的优缺点 甲醇酵母表达系统操作原理 甲醇酵母系统高效表达影响因素与对策 甲醇酵母表达系统的应用
1、甲醇酵母与甲醇氧化酶启动子
甲醇酵母(methylotrophic yeast) 指可利用甲醇作单一碳源的一类酵母。 毕赤酵母(Pichia pastoris) 汉森酵母(Hansenula ploymorpha) 假丝酵母(Candia boidinii)
2、甲醇酵母表达系统的优缺点
A、表达水平高(最高水平的系统) B、产物可翻译后修饰:糖基化、磷酸化、酰脂化 C、过糖基化程度比酿酒酵母少(8-15个vs100-150 个甘露糖) D、产物可正确折叠和高效分泌(最高分泌表达系统) E、可利用简单无机盐培养基高密度发酵,生物量大。 F、实验室和工业操作简单 G、不能满足结构要求严格的糖基化
2)基因剂量
外源基因表达存在基因剂量效应 筛选多拷贝整合子
载体引入G418/Zeocin抗性标记,整合子拷贝数 与抗性成正相关,采用高G418/Zeocin抗性转化子。
体外串联多个表达盒,直接获多拷贝整合子 采用YRp型载体稳定化技术获高拷贝整合子 构建高拷贝整合型表达载体
3)整合位点
外源基因表达盒整合于AOX/MOX或标记基因处,均 可高效表达
GAL80
GAL4
UAS
GAL1
GAL7
GAL10
A、 GAL1、GAL7和 GAL10基因连锁在一起,独立转录,共同调控 B、GAL4产物与UAS结合,促进转录;GAL80产物抑制GAL4产物的活性,阻遏转录
C、野生型GAL4表达水平低,产物活性可被GLAL80产物完全抑制,半乳糖诱导效果差
2)半乳糖激酶启动子(GAL1)
B、PHO4基因编码产物是PHO5启动子的正调控因子
C、pho4TS 温度敏感,35℃时失活,PHO5关闭 D、pho4TS-PHO5启动子通过降温(23℃)诱导表达
2、酿酒酵母分泌系统
酿酒酵母系统常用分泌信号肽来源:
性结合因子:MF-α
酸性磷酸酯酶:PHO5
蔗糖酶:SUC2 杀手毒素因子:KIL
宿主株:GS115、KM71 可插入位点: 5’AOX1 3’AOX1 TT 转化子: GS115:His+Mut+ KM71:His+Muts
2)HIS4位点插入
宿主株:GS115、KM71
可插入位点: 5’His4 3’His4
转化子: GS115:His+Mut+
KM71:His+Muts
3) 多基因插入事件(串联整合)
B、缺陷型标记基因:Leu2d 提高选择压力
C、抗性标记;neo
提高选择压力
甲醇酵母系统胞内表达载体
需要带入ATG
表达载体类型
单位点
甲醇酵母系统胞内表达载体
需要带入ATG
多位点
表达载体类型
甲醇酵母系统分泌表达载体
信号肽:PHO1
甲醇酵母系统分泌表达载体