纤维素的结晶结构
纳米纤维素结构

纳米纤维素结构
纳米纤维素是纤维素的纳米结构,是由纤维素分子组成的纳米晶体。
纤维素是一种天然聚合物,它是植物细胞壁的主要成分,也存在于一些藻类和真菌中。
纤维素是由葡萄糖分子通过β-1,4-糖苷键连接而成的线性多糖。
纳米纤维素的结构可以简单描述如下:
1. 分子结构:纳米纤维素由许多纤维素分子组成,每个纤维素分子是由许多葡萄糖分子连接而成。
葡萄糖分子之间通过β-1,4-糖苷键连接成线性链状结构。
2. 微观结构:纤维素分子可以自组装形成纤维状的纳米晶体。
这些纳米晶体具有纤维状的形态,通常呈现出直径约为数十纳米,长度可达数百纳米的纤维状结构。
3. 结晶结构:纳米纤维素的晶体结构是由多个纤维素分子通过氢键相互作用形成的。
这种结晶结构赋予纳米纤维素良好的力学性能和稳定性。
4. 纳米级细节:纳米纤维素的纤维形态使得其比表面积非常大,这意味着在相对较小的质量下,具有大量可用表面。
这些特性赋予纳米纤维素许多独特的特性和潜在应用。
纳米纤维素由于其独特的结构和性质,在生物医学、纳米技术、材料科学等领域有着广泛的应用。
它可以用于制备纳米纤维素膜、纳米纤维素凝胶、纳米纤维素纤维等材料,用于药物传递、组织工程、纳米复合材料等应用。
纤维素的结构

纤维素的结构引言纤维素(cellulose)是一种天然聚合物,它是植物细胞壁的主要成分,也是地球上最常见的有机化合物之一。
纤维素的结构不仅具有重要的生物学功能,而且在工业上有着广泛的应用价值。
本文将深入探讨纤维素的结构特点,包括化学组成、分子结构、晶体结构等方面的内容。
化学组成纤维素的化学式为(C6H10O5)n,其中n代表纤维素分子中重复单元的数量,可以是很大的一个数。
纤维素由葡萄糖分子通过β-1,4-糖苷键连接而成,因此纤维素可以看作是由许多葡萄糖分子组成的长链聚合物。
分子结构纤维素分子的结构比较复杂,由于葡萄糖分子通过β-1,4-糖苷键连接,使得纤维素分子呈现出直链的结构。
纤维素分子中的葡萄糖单元可以同时在链的不同位置上水解,因此纤维素分子具有较高的反应性。
纤维素分子的分子量较大,通常在几万到几十万之间。
纤维素的分子量与纤维素的来源有关,不同的植物纤维素具有不同的分子量分布。
晶体结构纤维素的晶体结构是纤维素研究的重要内容之一。
纤维素在自然界中以纤维素微纤维的形式存在,这些微纤维进一步结合形成纤维素纤锥,最终形成纤维素晶体。
纤维素晶体的晶格结构较为复杂,包含有多种晶体面。
其中最具有代表性的是纤维素I和纤维素II晶体。
纤维素I晶体是最常见的纤维素晶体形态,其晶体结构由两层纤维素链平行排列而成。
纤维素II晶体是较不常见的一种形态,其晶体结构由三层纤维素链交叉排列而成。
纤维素晶体具有很高的结晶度和强度,这使得纤维素在工业上具有广泛的应用。
纤维素的晶体结构还影响了纤维素的物理化学性质,如吸水性、热稳定性等。
分子间作用力纤维素分子之间通过多种分子间作用力相互吸引和排斥。
这些分子间作用力包括静电相互作用、范德华力、氢键等。
静电相互作用是纤维素分子间作用力的一种主要形式,纤维素分子中含有大量的羟基,这些羟基带有部分电荷,从而形成静电相互作用。
范德华力是一种瞬时极化引起的作用力,也是纤维素分子间相互吸引的重要力量。
纤维素结构

纤维素的结构引言纤维素是地球上存在的最丰富的可再生有机资源, 在高等植物、细菌、动物、海藻等生物中广泛存在, 每年总量有几百亿吨, 具有巨大的经济开发价值[1]。
五十年代至六十年代,由于合成高分子材料的兴起,纤维素资源的开发研究受到极大的影响。
七十年代初期,由于国际上出现了石油危机,这种曾被忽视的可更新资源又再次被重视起来.能否利用这些丰富的可再生资源是解决未来能源问题的关键因素。
因此,世界各国都很重视纤维素的研究与开发[2]。
纤维素结构是纤维素性能研究及应用的基础,本文就纤维素的化学剂物理结构进行了概述。
1纤维素的化学结构纤维素的元素组成为:C=44.44%,H=6.17%,O=49.39%, 其化学实验式(C 6H 10O 5)n (n 为聚合度,一般高等植物纤维素的聚合度为7000—150000)[3]纤维素大分子的基环是脱水葡萄糖,其分子式为(C 6H 10O 5)。
纤维素的化学结构是由D-吡喃葡萄糖环彼此以β- 1, 4-糖苷键以C1椅式构象联结而成的线形高分子化合物[4],其结构表达式如图1所示。
非还原端 纤维二糖 还原端图1 纤维素链结构除两端的葡萄糖基外,每个葡萄糖基上都有三个游离羟基,分别位于C 2、C 3和C 6位上,所以纤维素的分子可以表示为[[C 6H 7O 2(OH)3]n,其中C 2和C 3位上为仲醇羟基,C 6位上为伯醇羟基,他们的反应能力不同,对纤维素的性质具有重要影响,如纤维素的酯化、醚化、氧化和接枝共聚,以及纤维素之间的分子间氢键作用,纤维素的溶胀与水解都与纤维素的羟基有关。
纤维素大分子两端的葡萄糖末端基,其结构和性质不同,一端的葡萄糖末端基在C4上存在一个苷羟基,此羟基的氢原子易转移,与基环上的氧原子结合,使氧环结构转变为开链式结构,在C1处形成醛基,具有潜在还原性,固有隐形醛基之称。
左端的葡萄糖末端为非还原性的,由于纤维素的每一个分子链一端是还原性,另一端是非还原性,所以纤维素分子具有极性和方向性。
湿热-压缩处理木材的纤维素晶体结构变化

湿热-压缩处理木材的纤维素晶体结构变化尹江苹;郭娟;赵广杰;殷亚方【摘要】分别采用不同径向压缩率(25%和50%)并结合不同蒸汽处理温度(140、160 ℃和180℃)对人工林杉木(Cunninghamia lanceolata)进行湿热-压缩处理,然后利用X-射线衍射法(XRD)研究了湿热-压缩处理对木材纤维素微纤丝角、结晶度及微晶宽的影响规律.结果表明:湿热-压缩处理木材纤维素微纤丝角无明显变化,结晶度和微晶宽均增大.结晶度增幅介于28%~38%,不同压缩率和蒸汽处理温度对纤维素结晶度影响无显著差异.随着蒸汽处理温度的增加,微晶宽逐渐增大,在180℃蒸汽处理条件下达到最大值.%Compression combined with steam treatment to Chinese fir (Cunninghamia lanceolata) was carried out with different radial compression (compression ratio of 25% and 50%) in combination with different steam treatment temperatures (140,160 ℃ and 180 ℃).The variation of cellulose microfibril angle,crystallinity and crystallite width were characterized by X-ray diffraction(XRD).The results indicated that cellulose microfibril angle had no significant changes,however crystallinity and crystallite width were increased after CS-treatment.About 28%~38%increase of crystallinity was observed.Different compression ratios and steam treatment temperatures did not affect cellulose crystallinity significantly.With the increase of compression ratios,crystallite thickness increased gradually,the maximum value could be obtained when at 180 ℃.【期刊名称】《林产工业》【年(卷),期】2017(044)007【总页数】5页(P10-14)【关键词】杉木;湿热-压缩处理;纤维素;晶体结构【作者】尹江苹;郭娟;赵广杰;殷亚方【作者单位】中国林业科学研究院木材工业研究所;;;中国林业科学研究院木材工业研究所【正文语种】中文【中图分类】S781.7由于天然林资源的短缺及人类保护意识的增强,人工林木材资源将逐步成为木材资源战略储备中的重要组成。
醋酸纤维素的结晶结构与热性能

b c u e o t mp r c rsalt e a s fisi ef tc tl e;o h t e a d, ga sta sto fCDA smoe e ie t a d i Tg e y i n t e oh rh n ls rn i n o i wa r vd n , n t s
第2 9卷 第 1 O期
20 0 8年 l 0月
纺
织
学
报
V0 . 9 No. O 12 1
J u n lo xie Re e rh o r a fTe t sa c l
Oc . 2 O8 t 0
文 章编 号 :2 3 7 1 2 0 )00 1—5 0 5 — 2 (0 8 1 .0 20 9
醋 酸 纤 维 素 的 结 晶 结 构 1 中原 工 学 院 纺 织学 院 , 南 郑 州 河 400 ; . 华 大 学 纺 织 学 院 , 海 507 2 东 上 2 12 ) 060
摘
要
研究三醋酸纤维素(T ) C A 和二 醋 酸 纤 维 素 ( D 的 结 晶结 构 和 热 学 性 能 。纤 维 素 乙酰 化 后 形 成 C A, C A) T 其
晶型 发生 变 化 , 晶度 降 低 ; C A 水 解 后 得 到 的 C A 无 定 形 占明 显 优 势 , 丝 过 程 中 的 取 向 又 会 进 一 步 降 低 结 而 T D , 纺
纤维素知识

互穿聚合物网络(IPN)天然纤维素包括细菌纤维素、海藻和高等植物(如棉花、苎麻、木材等)均属于纤维素I型。
纤维素I分子链在晶胞内是平行堆砌的,纤维素II是纤维素I经由溶液中再生(regeneration)或经丝光处理(mercerization)得到的结晶变体,是工业上使用最多的纤维素形式。
纤维素II 与纤维素I有很大的不同,它是由两条分子链组成的单斜晶胞,属于反平行链的堆砌。
纤维素是一种β-(1-4)-D-糖苷键连接的线型高聚物,由X 射线衍射发现存在四种结晶形态,即纤维素Ⅰ、Ⅱ、Ⅲ和Ⅳ,不同晶型纤维素的C1,C4 和C6 的化学位移具有明显的差别,这种差别可能是因为不同晶型纤维素的链构象转变或晶体堆砌对吡喃葡萄糖单元C4 和C6 的影响差异造成的.基于在非晶区的链段运动显示窄谱线,而晶区的刚性链以及分布在非晶区的刚性链产生宽谱线,因此可以利用宽、窄谱线的峰面积(S b 和S n)求取结晶度χ c纤维素I和纤维素II在C6上的差别就是因为吡喃葡萄糖单元C6位羟基的构象不同,纤维素I为t - g构象,纤维素II和无定型纤维素则为g - t构象。
天然纤维素I也存在两种不同的晶体结构,即纤维素Iα和Iβ。
13C NMR谱指出它们之间最大的差别在C1的化学位移上,I α为单峰,Iβ为双峰。
高分子链构象参数:无扰尺寸(A):A 值愈小,高分子链愈柔顺。
A 值只取决于高分子的近程结构,与高聚物的分子量无关。
空间位阻参数(σ):空间位阻参数是指由于高分子链的内旋转受阻而导致分子尺寸增大程度的量度,其值愈小,高分子链愈柔顺Flory 极限特征比(C∞):Flory 极限特征比是指高分子链由于键角限制和空间位阻造成分子链伸展的程度,。
一般来说,C ∞愈小,链愈柔顺。
合成的柔顺性高聚物的C值在5~7 范围内,而天然高分子多数高于此范围。
持续长度(q):持续长度q 广义为高分子链在第一个键方向上的投影,它表征分子链的支撑能力。
纤维素的结构及性质
一.结构纤维素是一种重要的多糖,它是植物细胞支撑物质的材料,是自然界最非丰富的生物质资源。
在我们的提取对象-农作物秸秆中的含量达到450-460g/kg。
纤维素的结构确定为β—D-葡萄糖单元经β-(1→4)苷键连接而成的直链多聚体,其结构中没有分支。
纤维素的化学式:C6H10O5化学结构的实验分子式为(C6H10O5)n早在20世纪20年代,就证明了纤维素由纯的脱水D-葡萄糖的重复单元所组成,也已证明重复单元是纤维二糖。
纤维素中碳、氢、氧三种元素的比例是:碳含量为44。
44%,氢含量为6.17%,氧含量为49.39%。
一般认为纤维素分子约由8000~12000个左右的葡萄糖残基所构成.O OOOOOOOO1→4)苷键β-D-葡萄糖纤维素分子的部分结构(碳上所连羟基和氢省略)二.天然纤维素的原料的特征做为陆生植物的骨架材料,亿万年的长期历史进化使植物纤维具有非常强的自我保护功能。
其三类主要成分-纤维素、半纤维素和木质素本身均为具有复杂空间结构的高分子化合物,它们相互结合形成复杂的超分子化合物,并进一步形成各种各样的植物细胞壁结构。
纤维素分子规则排列、聚集成束,由此决定了细胞壁的构架,在纤丝构架之间充满了半纤维素和木质素。
天然纤维素被有效利用的最大障碍是它被难以降解的木质素所包被。
纤维素和半纤维素或木质素分子之间的结合主要依赖于氢键,半纤维素和木质素之间除了氢键外还存在着化学健的结合,致使半纤维素和木质素之间的化学健结合主要在半纤维素分子支链上的半乳糖基和阿拉伯糖基与木质素之间。
表:植物细胞壁中纤维素、半纤维素、和木质素的结构和化学组成项目纤维素木质素半纤维素结构单元吡喃型D-葡萄糖基G、S、H D-木糖、苷露糖、L-阿拉伯糖、半乳糖、葡萄糖醛酸结构单元间连接键β—1,4—糖苷键多种醚键和C—C键,主要是β-O-4型醚键主链大多为β-1,4-糖苷键、支链为β-1,2-糖苷键、β-1,3—糖苷键、β-1,6-糖苷键聚合度几百到几万4000 200以下聚合物β—1,4-葡聚糖G木质素、GS木质素、GSH木质素木聚糖类、半乳糖葡萄糖苷露聚糖、葡萄糖甘露聚糖结构由结晶区和无定型区两相组成立体线性分子α不定型的、非均一的、非线性的三维立体聚合物有少量结晶区的空间结构不均一的分子,大多为无定型三类成分之间的连接氢键与半纤维素之间有化学健作用与木质素之间有化学健作用天然纤维素原料除上述三大类组分外,尚含有少量的果胶、含氮化合物和无机物成分。
纤维素(三) - 纤维素(三)
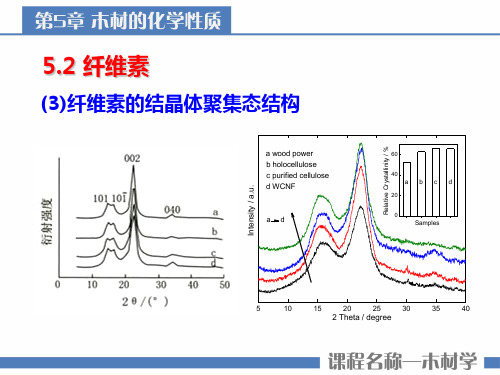
(3)纤维素的结晶体聚集态结构
a wood power b holocellulose c purified cellulose d WCNF
ad
60
40 abcd
20
0 Samples
Intensity / a.u.
Relative Crystallinity / %
5
10
15
20
25
5.2 纤维素
(4)纤维素的物理性质
表面电化学性质:
①羟基和醛基可使纤维素表 面在水溶剂中带负电;
②正电子由于热运动结果在 纤维表面由近而远有一浓 度分布。
(a)离子层 (b)吸附层 (d) 扩散层
5.2 纤维素
(4)纤维素的物理性质
表面电化学性质:
在湿法纤维板制造工艺中,为了减少纤维的吸湿,一 般在纤维板的浆料中施加石蜡乳液进行阻水处理。但 石蜡微粒带有负电荷,于浆料中加入硫酸铝作沉淀剂 ,使石蜡留存在纤维表面上,经处理的浆料制成的纤 维板,防水性能和体积稳定性均有所改善,提高产品 质量。
5.2 纤维素
(5)纤维素的化学性质
纤维素的降解 氧化降解:
例:2,2,6,6-四甲基哌啶氧自由基(TEMPO)的选择性催化氧化
纤维素经氧化剂作用后,羟基氧化成醛基、酮基或 羧基,形成氧化纤维素。随着官能团的变化,纤维素 的聚合度也同时下降,发生氧化降解后,纤维素的机 械强度降低。
5.2 纤维素
5.2 纤维素
(5)纤维素的化学性质
纤维素的酯化
如醋酸酐与纤维素的-OH基作用
[C6H7O2(OH)3]n+3n(CH3CO)2O→ [C6H7O2(OCOCH3)3]n + 3nCH3COOH
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
:s与a之夹角
点阵基矢(原子间距)a
入射线单位矢量s0
0:s0与a之夹角
一维劳埃方程的导出
原子列中任意两相邻原子(A与B)散射线间光程差()为: =AM-BN=acos-acos0 散射线干涉一致加强的条件为=H,即: a(cos-cos0)=H H——任意整数
纤维素X与纤维素IV晶胞参数大致相等。
纤维素结晶变体的相互转化
+液体氨
蒸发
纤维素I
+H2O 100º C +NaOH 100º C +NaOH 20º C
NH3-纤维素I
≥200º C
纤维素ⅢI
Na-纤维素II
+NaOH
Na-纤维素III
+H2O +液体氨 +H2O 蒸发
纤维素IVI
纤维素II
NH3-纤维素II
CHINESE ACADEMY OF FORESTRY
纤维素结晶结构
组员:郭 飞 张毛毛 周贤武 温留来 张龙飞
2012-12-04
1
3.纤维素的物理结构
3.1 纤维素的结晶结构 晶体的基本概念
晶体:物质内部质点(原子、分子、离子)呈规律排列的固体, 为“空间格子”状。 晶胞:晶体的最小重复单位。可以用3个晶轴的长度a、b、c 及其夹角α 、β 、γ 6个晶胞参数来描述。
或
a· 0)=H (s-s b· 0)=K (s-s c· 0)=L (s-s 三维晶体若要产生衍射,必须同时满足上述三个方程
Scherrer公式
Scherrer公式用于样品晶粒尺寸的计算
D=K·λ /(Bhkl ·cosθ)
其中,D为沿垂直于晶面(hkl)方向的晶粒直径,
k为Scherrer常数(通常为0.89), λ为入射
素和木质素,因此引入相对结晶度的概念。一般有两种方
法:
纤维素相对结晶度
(1)找出非结晶区的衍射曲线
(2)Segal法和Turley法
纤维素I结晶度计算
CrI(%)=(I002-Iam)/ I002 ×100
•Iam — 2 θ为18º 附近无定形区散 射强度 •I002 —(002)面的衍射强度 纤维素II结晶度计算
晶面和晶面指数
纤维素单元细胞的结晶变体
固态下的纤维素存在5种结晶变体,即天然纤维素I、人造纤 维素II、纤维素III、纤维素IV和纤维素X。5种结晶变体各有 不同的晶胞结构,晶胞参数如下: a 纤维素I 纤维素II 纤维素III 纤维素IV 8.35Å 8.10Å 7.74Å 8.11Å b 10.3Å 10.3Å 10.3Å 10.3Å c 7.9Å 9.1Å 9.9Å 7.9Å β 84º 62º 58º 90º
比可见光镜面反射安排实验,用X射线照射岩盐
(NaCl),并依据实验结果导出布拉格方程。
一、布拉格方程
散射角2 :入射线方向与 散射线方向之间的夹角。 选择反射:当X射线以某 些角度入射时,记录到 反射线,其它角度入射, 则无反射。 如:以Cu K射线照射 NaCl表面,当=15和 =32时记录到反射线。
1.布拉格实验
设入射线与反射面之夹角为,称掠射角或布拉格角,则按反射定律, 反射线与反射面之夹角也应为。
2.布拉格方程的导出
考虑到: ①晶体结构的周期性,可将晶体视为由许多相互平行且晶 面间距(d)相等的原子面组成; ②X射线具有穿透性,可照射到晶体的各个原子面上; ③光源及记录装置至样品的距离比 d 数量级大得多,故入 射线与反射线均可视为平行光。
结晶区是连续的缨状原纤。由许多长链分子组成。(原纤即纤丝)
中的晶区变得很短的情况。
3.折叠链结晶结构理论 (fringed-micelle with chain-folding)
纤维素大分子有呈折叠链状的可能,这些折
叠链状的线型纤维素大分子仍然能形成结晶
结构,即所谓折叠链结晶,或所谓片晶。人 们认为片晶就如同缨状微胞结构中的微胞;
相对结晶度衍射示意图(王欣 香根草构造与基本特性研究)
X-射线衍射法测定纤维素相对结晶度案例-1
球磨时间增加,结晶度下降
X-射线衍射法测定纤维素相对结晶度案例-1
纤维素结晶区与非结晶区的关系
纤维素纤维中纤维素的聚集态是很复杂的。 一相结构理论没有得到公认。
现在较普遍承认的是两相结构理论,即纤维素纤维中的纤
维素是以结晶相和无定形相共存的。
纤维素结晶区与非结晶区的关系 1.缨状微胞理论
长链分子间的规整排列构成结晶微胞。而伸出的无 规则排列的分子成为缨状须从。故称为缨状微胞理 论(fringed micelle theory)。 (1)纤维素大分子链不在一个微胞内终 止,而是贯穿了一个以上的微胞(晶区) 和微胞间物质(非晶区),分子长度与 微胞长度无一定关系,晶区和非晶区无 明显的界面。 (2)纤维素大分子从晶区逸出以缨状的形
CrI%=(I101-Iam)/ I101 × 100 Iam — 2θ为15º 附近无定形区散射 强度 •I101—(101)面的衍射强度
Turley法
将样品木粉在室温下 压成薄片,然后做成 2θ强度曲线,样品扫 描范围为3°~ 40°( 2θ)角, 在扫 描曲线上2θ = 22°附近(002)衍 射的极大峰值 ( I002 ) , 2θ= 18°附近有一极小 值( Iam ) 。
布拉格将X射线的“选择反射”解释为:
入射的平行光照射到晶体中各平行原子面上,各原子面 各自产生的相互平行的反射线间的干涉作用导致了“选 择反射”的结果。
布拉格方程的导出
设一束平行的X射线(波长)以 角照射到晶体中晶面指 数为(hkl)的各原子面上,各原子面产生反射。 任选两相邻面(A1与A2),反射线光程差=ML+LN=2dsin ; 干涉一致加强的条件为=n,即 2dsin=n 式中:n——任意整数,称反射级数,d为(hkl)晶面间距, 即dhkl。
a(cos-cos0)=H表达了单一原子列衍射线方向()
与入射线波长()及方向(0)和点阵常数的相互关系,
称为一维劳埃方程。 亦可写为 : a· 0)=H (s-s
2. 二维劳埃方程
a(cos-cos0)=H b(cos-cos0)=K
0及0——s0与a及b的夹角 及——s与a及b的夹角
≥200º C
纤维素ⅢII
纤维素IVII
纤维素 I的晶胞特征
纤维素I结晶格子是单斜晶体,即具有3条不同长度的轴和一
个非90°的夹角。纤维素I的结构,介绍如下2种模型:
Meyer-Misch 模型 ① 晶胞参数: a=8.35Å,b=10.3Å ,c=7.9Å β=84º 纤维素分子链只占据结晶单元 的4个角和中轴,而每个角上的链为4 个相邻单位晶胞所共有,即每个晶胞 只含2个(4*1/4+1)链单位。 ② 中间链和位于角上的链走向相反,轴 向高度差半个葡萄糖基。b轴的长度 是纤维二糖的长度,这些链围绕着纵 轴扭转180°。
纤维素 II的晶胞特征
① 属于单斜晶系,晶胞参数平均值: a=7.93Å,b=9.18Å , c=10.34Å,γ=117.31º ② 相邻分子链是反向平行。 ③ 角链上的伯羟基为gt位,链的方向向上,中心链上的伯羟 基为tg位,链的方向向下。 ④ 中心链相对于角链在纤维轴c的方向上相互错开0.216c。 ⑤ 纤维素 II中形成的氢键网较纤维素 I中复杂。前者比后者堆 砌较为紧密,在热力学上更稳定。
Blackwell模型
① 晶胞参数: a=16.34Å,b=15.72Å,c=10.38Å,β =97.0º ② 纤维素分子链占据结晶单元的 4个角和中轴。(这种晶胞含8 条分子链横截面) ③ 中间链和位于角上的链是沿同 一方向的平行链,中间链在高 度上与位于角上的链半个葡萄 糖基。 ④ 链分子的薄片平行于ac面,所有 的伯羟基均为tg构象(见第38 页)。 ⑤ a轴方向形成分子内氢键,还 形成分子间氢键。
相 对 强 度
X射线管结构
X-射线衍射法
X-射线衍射的基本原理
晶体是由原子或原子基团等按照一定规律在空间内有 规则排列而构成的固体。当它被X射线照射后,各个原
子散射X射线。这些散射线符合相干波的条件,因而产
生干涉现象பைடு நூலகம்所谓X射线衍射,实质上就是研究这些散
射波的干涉。衍射线就是经过相互干涉而加强的大量散
交叉极化魔角旋转核磁共振法
拉曼光谱
X-射线衍射法
X-射线的产生与X射线
当高速运动的电子撞击到一个金属靶上时,靶面上被电子
撞击的部位就产生电磁波辐射,其中一部分为X射线。在
管电压没超过某一数值Vk(激发电压)时,只有连续射线 谱产生。超过Vk时,若干强度很高的特征谱线叠加在连续
谱线上,为特征X射线谱。
式进入非晶区,由高度结晶的有序区至
完全无结晶的无序区是连续过渡的。
2.缨状基微纤维理论(fringed-fibril theory)
(1)又称“缨状原纤结构理论”,放弃了晶区是微胞的假设, . (2)由于巨分子聚集过程中的缠结和局部无序,晶区中的分子 不在同一位置上逸出,也不肯无限地结合在同一结晶原纤中, 而可在晶区不同的部位上离开,造成原纤中晶区的弯曲、扭 变和分叉,所以原纤在横向和长向上都 可不断地分裂和重建,构成网络组织的 晶区和非晶区。 认为缨状微胞理论是长的缨状原纤的极 限情况,即当结晶期间成核频繁,原纤
射线所组成的射线。
X-射线衍射法
衍射花样和晶体结构的关系:
每种晶体所产生的衍射花样都反映出晶体内部的原子
分布规律。 衍射花样的特征由两个方面组成:一是衍射线在空间 的分布规律,由晶胞的大小、形状和位向绝对;二是 衍射线的强度,取决于原子在晶胞中的位置、数量和 种类。
晶胞大小的研究方法