表观遗传学 第三章 组蛋白修饰 ppt课件
表观遗传学教学课件

04
表观遗传学研究方法
基因组学技术
基因组测序
通过全基因组测序技术,可以检测基因组中的变异和表观遗传修饰,了解基因表达的调 控机制。
甲基化测序
甲基化测序技术可以检测基因组中DNA甲基化的水平,研究甲基化与基因表达的关系。
生物信息学分析
数据挖掘
利用生物信息学方法对大规模基因组 数据进行挖掘,寻找表观遗传修饰与 基因表达之间的关联。
详细描述
非编码RNA在表观遗传学中发挥重要作用, 它们通过与mRNA相互作用,影响基因表达 的转录和转录后水平。非编码RNA的异常表 达与多种疾病的发生和发展密切相关。
组蛋白修饰
总结词
组蛋白修饰是指组蛋白上的化学基团, 如乙酰化、甲基化和磷酸化等。
VS
详细描述
组蛋白修饰能够影响染色质的结构和基因 表达,与细胞分化、发育和肿瘤形成等生 物学过程密切相关。组蛋白修饰的异常与 多种疾病的发生和发展密切相关。
80%
药物研发
表观遗传学研究有助于发现新型 药物靶点,推动药物研发的创新 和进步。
表观遗传学面临的挑战与问题
技术难题
表观遗传学研究涉及多种复杂技 术,如高通量测序、染色质免疫 沉淀等,技术难度较大,需要专 业人员操作。
数据解读与分析
表观遗传学研究产生大量数据, 如何准确解读和分析这些数据是 一个挑战。需要发展新的数据分 析方法和算法。
个体化治疗
表观遗传学研究有助于实现个 体化治疗,即根据患者的表观 遗传学特征,制定个性化的治 疗方案。例如,针对特定基因 的靶向治疗等。
疾病预防
表观遗传学研究还有助于疾病 的预防。例如,通过调整饮食 和生活方式等,可以改变个体 的表观遗传学特征,从而预防 某些疾病的发生。
表观遗传学(共49张PPT)
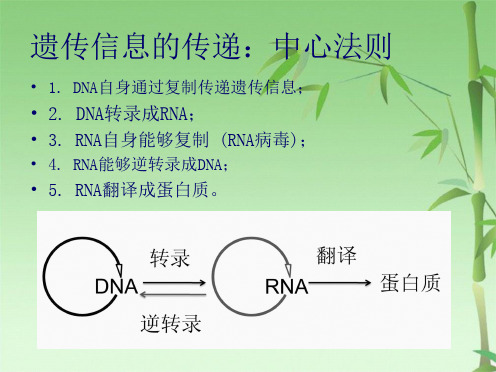
• 1. DNA自身通过复制传递遗传信息;
• 2. DNA转录成RNA; • 3. RNA自身能够复制 (RNA病毒);
• 4. RNA能够逆转录成DNA;
• 5. RNA翻译成蛋白质。
• 1939年,生物学家 Conrad Hal Waddington首先在《现代遗传学导论》
微小RNA(microRNA ,miRNA—单链)。
• RNA干扰(RNAi):是通过小RNA分子在mRNA水平上介导mRNA 的降解诱导特异性序列基因沉默的过程。
• 诱导染色质结构的改变,决定着细胞的分化命运,还对 外源的核酸序列有降解作用以保护本身的基因组。
21
2.长链非编码RNA (long noncoding RNA, lncRNA)
DXPas34 长度超过200bp;
DNA甲基化状态的保
持
11
• (一)DNMTs(DNA methyltransferases)
DNA甲基转移酶 结构特点:
-NH2末端调节结构域,介导胞核定位,调节与其他蛋白相互 作用。DNMT2无。
-COOH末端催化结构域,参与DNA甲基转移反应。 • 1.DNMT1
20
• 三、其他表观遗传过程
• (一)非编码RNA的表观遗传学
• 非编码RNA(non-protein-coding RNA,ncRNA)
• tRNA,rRNA;短链非编码RNA,长链非编码RNA。
• 短链RNA(又称小RNA),小干涉RNA(short interfering RNA ,siRNA—双链) 和
S-腺苷甲硫氨酸: S-adenosylmethionine,SAM S-腺苷同型半胱氨酸:S-adenosylhomocysteine,SAH
组蛋白修饰及其功能(乙酰化,甲基化,磷酸化等)-于凯讲解学习

表观遗传学(epigentics)是研究不改变DNA序列而由于其外 部修饰引起的基因开放与否的学科,涉及的主要机制有DNA甲基 化、组蛋白修饰、基因印记、RNA干扰等。其中研究得最多是 DNA甲基化和组蛋白乙酰化、组蛋白甲基化,这些修饰与活化或 失活染色质的结构形成相关。
染色质是由许多核小体组成的,大部分真核生物中有5种富含 碱性氨基酸的组蛋白,即H1,H2A,H2B,H3和H4。H2A,H2B, H3和H4各2个分子构成的8聚体是核小体的核心部分,H1的作用是 与线形 DNA结合以帮助后者形成高级结构。
组蛋白翻译完成后,其氨基尾巴会发生多种共价修饰,如乙 酰化、甲基化、磷酸化,泛素化和ADP核糖基化等,这些修饰都 是可逆性修饰,这些修饰共同构成了“组蛋白密码”。
1. 组蛋白乙酰化
核心组蛋白乙酰化反应多发生在核心组蛋白 N端碱性氨基 酸集中区的特定 Lys 残基。组蛋白乙酰化由组蛋白乙酰转移酶 (histone acetyltransferase,HAT)和组蛋白去乙酰化酶(histone deacetylase,HDAC)协调进行。HAT通过将乙酰辅酶 A 的乙酰 基转移到 Lys 的NH+,中和掉一个正电荷。 HDAC使组蛋白去乙 酰化,与带负电荷的DNA紧密结合,染色质致密卷曲,基因的 转录受到抑制。
2. 组蛋白的甲基化
组蛋白甲基化是由组蛋白甲基化转移酶(histone methyl transferase,HMT)完成的。甲基化可发生在组 蛋白的赖氨酸和精氨酸残基上,而且赖氨酸残基能够 发生单、双、三甲基化,而精氨酸残基能够单、双甲 基化,这些不同程度的甲基化极大地增加了组蛋白修 饰和调节基因表达的复杂性。
局部乙酰化举例
当DNA与核小体尚未解开缠绕时,转录激活因子如糖皮质激素受体可以和DNA上相应 的反应元件(GRE)结合。当结合至GRE之后,糖皮质激素募集共激活因子如CBP到染色 体上的靶转录基因区。此时,共激活因子利用HAT活性使得结合在DNA启动子区域的核心 组蛋白乙酰化,进而使DNA与组蛋白结合减弱,核小体释放,转录因子和RNA聚合酶可以 与DNA上特异的启动子结合,启动靶基因的转录。
表观遗传学——甲基化,组蛋白修饰

表观遗传学——甲基化,组蛋⽩修饰参考资料:1.2.3.1.什么是表观遗传学?举个例⼦:同卵双⽣的双胞胎个体,从遗传学⾓度说他们的DNA序列是⼀致的,但多种表型存在⼀些差异。
经典的孟德尔遗传定律和⽣物学表型之间还存在另外⼀层调控因素,即表观遗传。
表观遗传(Epigenetics)是指DNA序列未发⽣变化,但基因表达却发⽣了可遗传改变。
这种改变的特点:可遗传性;可逆性;没有DNA序列的变化。
可逆性:表观遗传的修饰⽅式可以在某些因素的条件下被去除。
这使得通过调控表观遗传来影响⽣物学性状称为可能。
表观遗传改变主要从四个层⾯调控基因表达(1)DNA甲基化:DNA共价结合甲基基团,使相同序列等位基因处于不同修饰状态;(2)组蛋⽩修饰:通过对结合DNA的组蛋⽩进⾏不同的化学修饰实现对基因表达的调控;(3)染⾊质重塑:通过改变染⾊质的空间构象实现对基因表达的调控;(4)⾮编码RNA的调控:RNA可通过某些机制实现对基因转录和转录后的调控。
2.DNA甲基化DNA序列上特定的碱基在DNA甲基转移酶(DNMT)的催化作⽤下,以S-腺苷甲硫氨酸(SAM)作为甲基供体,通过共价结合的⽅式获得⼀个甲基基团的化学修饰过程。
最常见能够被甲基化的碱基是胞嘧啶(C),此外腺嘌呤,鸟嘌呤也可以被甲基化。
下图是5甲基胞嘧啶。
在4位上是⼀个胺基,5位上没有其他基团的结合。
在SAM提供甲基的情况下,在DNMT(DNA甲基转移酶)的作⽤下,甲基从SAM转移到胞嘧啶的5位,成为了5甲基胞嘧啶。
DNA甲基转移酶根据序列的同源性和功能,真核⽣物DNA甲基化转移酶主要分为:Dnmt 1, Dnmt2 和Dnmt 3.Dnmt 1参与序列甲基化的维持; Dnmt 3主要作⽤是从头甲基化。
a图左边的序列通过Dnmt 3的作⽤转化为右边的序列,这两个序列的差别是,所有的C(互补链上)被甲基化,这是⼀种重头甲基化的⽅式。
b图中左边的序列其中⼀条链上C位点被甲基化,互补链上的C没有甲基化,可以在甲基化维持酶(Dnmt 1)的作⽤下可以使得另外⼀条⾮甲基化的链进⾏甲基化。
表观遗传学(研究生课件)

染色质重塑的研究方法
• 研究染色质重塑的方法包括遗传学方法、生物化学方法以及显 微镜技术等。遗传学方法包括基因敲除和转基因技术等,可以 用于研究染色质重塑酶和组蛋白修饰酶的功能。生物化学方法 包括蛋白质纯化和结晶化技术、质谱分析和代谢组学技术等, 可以用于研究染色质重塑酶和组蛋白修饰酶的相互作用和生物 化学性质。显微镜技术则可以用于观察染色质结构和动态变化。
基因组学方法
通过基因组学技术,研究非编码RNA的基因组位置、 序列和结构等信息。
转录组学方法
通过转录组学技术,研究非编码RNA的表达水平和转 录本信息。
蛋白质组学方法
通过蛋白质组学技术,研究非编码RNA对蛋白质表达 和功能的影响。
05
表观遗传学与疾病
表观遗传学与肿瘤
肿瘤表观遗传学
研究肿瘤发生发展过程中表观遗传机 制的改变,包括DNA甲基化、组蛋白 修饰和非编码RNA等。
表观遗传学的研究内容
总结词
表观遗传学的研究内容包括表观遗传修饰的机制、表观遗传与疾病的关系以及表观遗传修饰的干预策 略。
详细描述
表观遗传学研究DNA甲基化、组蛋白修饰和非编码RNA调控等机制,探讨这些修饰如何影响基因表达 和细胞功能。同时,研究表观遗传学与各种疾病的关系,包括癌症、神经退行性疾病和代谢性疾病等 。此外,还研究如何通过干预表观遗传修饰来治疗疾病。
表观遗传学的重要性
总结词
表观遗传学在理解生物学过程、疾病机制和治疗策略方面具有重要意义。
详细描述ห้องสมุดไป่ตู้
表观遗传学在理解细胞分化、胚胎发育和衰老等生物学过程中发挥关键作用。同时,表观遗传学与许多疾病的发 生和发展密切相关,为疾病的诊断和治疗提供了新的视角。此外,表观遗传修饰的可逆性为疾病治疗提供了潜在 的干预策略,有助于开发新的治疗方法和药物。
组蛋白修饰与表观遗传学

组蛋白修饰与表观遗传学在生物学中,基因组是由DNA组成的,而基因组的表达可能会因一些因素的影响而受到调控。
这些因素之一就是组蛋白修饰。
组蛋白修饰是一种通过改变组蛋白的状态来调节基因表达的方式。
本文将详细介绍组蛋白修饰的方式及其在表观遗传学中的作用。
组蛋白修饰的方式组蛋白是一种主要存在于细胞核中的蛋白质,它可以将DNA紧密地包裹起来,形成染色体。
在此基础上,组蛋白还有其他功能,例如:保护DNA免受化学和机械伤害、调控DNA复制和修复等。
组蛋白修饰的主要方式是通过化学修饰来改变组蛋白的状态。
这些化学修饰包括乙酰化、甲基化、磷酸化、泛素化和丝氨酸/苏氨酸磷酸化等。
这些化学修饰可以在组蛋白的N端、C端或其间的Lys和Arg等残基上进行。
乙酰化是最常见的一种组蛋白修饰方式之一,它通过在氨基或羟基侧链上添加乙酰基来改变组蛋白的状态。
当组蛋白乙酰化后,它会呈现出更开放的构象,使得DNA更容易被转录因子访问,同时还可以吸引一些辅助蛋白质来调控基因表达。
除了乙酰化,甲基化也是比较常见的一种组蛋白修饰方式。
它通过添加甲基基团来改变组蛋白的状态,这一过程会影响染色体的执行效率,例如抑制转录。
组蛋白修饰的作用组蛋白修饰在表观遗传学中有着极其重要的作用。
表观遗传学指的是不涉及DNA序列本身的遗传学现象。
基因组的表达在常态下往往是受到组蛋白修饰的影响而被调节的。
一方面,组蛋白修饰可以调节染色体的可及性。
当组蛋白乙酰化或磷酸化后,组蛋白的整体紧密度会改变,使得DNA更容易被其他蛋白质访问,这对于DNA复制,转录和修复等过程都是必要的。
另一方面,组蛋白修饰可以影响染色体上基因的表达。
以乙酰化为例,乙酰化可以使得基因的转录水平上升,从而改变细胞的表现和行为。
甲基化则是比较特殊的一种组蛋白修饰方式,它更多地与静默基因有关,静默基因的存在意味着某种基因的表达在特定的情况下被抑制,这个状态可以维持数个细胞周期之久。
组蛋白修饰与疾病近年来,研究人员发现很多疾病与组蛋白修饰之间存在一定的关联。
表观遗传学课件(带目录)
表观遗传学课件一、引言表观遗传学是研究基因表达调控机制的一门学科,它涉及到基因序列不发生变化,但基因表达却发生了可遗传的改变。
这种调控机制对于生物体的生长发育、细胞分化、疾病发生等过程具有重要作用。
本文将对表观遗传学的基本概念、调控机制及其在疾病中的应用进行详细阐述。
二、表观遗传学的基本概念1.基因表达调控:基因表达调控是指生物体通过一系列机制,控制基因在特定时间和空间的表达水平。
基因表达调控是生物体生长发育、细胞分化、环境适应等生命现象的基础。
2.表观遗传修饰:表观遗传修饰是指在基因的DNA序列不发生改变的情况下,通过DNA甲基化、组蛋白修饰、染色质重塑等机制调控基因表达的过程。
3.表观遗传学的研究内容:表观遗传学主要研究基因表达调控的分子机制,包括DNA甲基化、组蛋白修饰、染色质重塑、非编码RNA调控等。
三、表观遗传学的调控机制1.DNA甲基化:DNA甲基化是指在DNA甲基转移酶的催化下,将甲基基团转移至DNA分子的过程。
DNA甲基化通常发生在基因的启动子区域,抑制基因表达。
2.组蛋白修饰:组蛋白修饰是指在组蛋白分子上发生的一系列化学修饰,如乙酰化、磷酸化、甲基化等。
这些修饰可以改变组蛋白与DNA的结合状态,从而调控基因表达。
3.染色质重塑:染色质重塑是指染色质结构发生变化,使基因的表达状态发生改变的过程。
染色质重塑可以通过改变核小体结构、DNA甲基化、组蛋白修饰等方式实现。
4.非编码RNA调控:非编码RNA是指不具有编码蛋白质功能的RNA分子,包括miRNA、lncRNA、circRNA等。
这些RNA分子可以通过与mRNA结合、调控转录因子活性等方式调控基因表达。
四、表观遗传学在疾病中的应用1.癌症:表观遗传学在癌症研究中的应用主要涉及肿瘤发生、发展和治疗。
研究发现,癌细胞的表观遗传修饰模式发生改变,导致肿瘤相关基因的表达异常。
通过研究这些表观遗传修饰,可以为癌症的早期诊断、预后评估和治疗提供新靶点。
《表观遗传学》PPT课件-2024鲜版
染色质构象捕获技 术
结合高通量测序和生物信息学分析,研究非编码RNA与染 色质构象的关系及其对基因表达的调控作用。
2024/3/27
26
07
表观遗传学前沿与展望
2024/3/27
27
表观遗传学领域的研究热点
表观遗传学定义
研究基因表达或细胞表现型的变化, 而非DNA序列改变的科学。
发展历程
从经典遗传学到分子遗传学,再到表 观遗传学,人类对基因表达调控的认 识不断深入。
2024/3/27
4
表观遗传学与遗传学的关系
2024/3/27
遗传学
01
研究基因序列的遗传与变异规律。
表观遗传学
02
研究基因表达调控的规律,与遗传学相辅相成。
17
表观遗传与生物进化
2024/3/27
表观遗传变异与自然选择
生物体在应对环境压力时,可能通过表观遗传变异产生适应性表型。这些变异可以在不改变DNA序列的情 况下传递给后代,并在自然选择的作用下逐渐在种群中累积。
表观遗传与物种形成
在物种形成过程中,生殖隔离的形成是至关重要的。表观遗传机制可以在不影响DNA序列的情况下,导致 不同种群间基因表达的差异,进而促进生殖隔离的形成和物种的分化。
表观遗传与生物复杂性
生物体的复杂性不仅体现在基因组的多样性上,还体现在基因表达的精细调控上。表观遗传机制通过影响基 因表达的时空特异性和水平,为生物复杂性的产生和维持提供了重要的调控手段。
18
05
表观遗传与人类疾病
2024/3/27
19
肿瘤与表观遗传异常
遗传学:表观遗传学教学课件
基因表达模式
• 决定细胞类型的不是基因本身,而是基因 表达模式,通过细胞分裂来传递和稳定地 维持具有组织和细胞特异性的基因表达模 式对于整个机体的结构和功能协调是至关 重要的。
• 基因表达模式在细胞世代之间的可遗传性 并不依赖细胞内DNA的序列信息。
• 基因表达模式有表观遗传修饰决定。
Waddington's epigenetics
• CMT3 (CHROMOMETHYLASE3) – 5'-CHG-3' sites
• (H= A, C or T) • Interacts with histone mark
• CMT2 (CHROMOMETHYLASE3) – 5'-CHH-3' sites
DRM 1, DRM 2 (DOMAINS REARRANGED 1 and 2) - 5'-CHH-3' sites
Photo credit: DrL
Mosaicism: An Individual with Two Different Eye Colors
“Diego”
Mosaicism: An Individual Eye with Two Colors
Epigenetic programming in plants helps silence transposons and maintain centromere function
2、衰老
无论DNA甲基化水平增高还是减低,都与人的 衰老过程相关。
3、免疫紊乱 在狼疮病人的T细胞中,甲基转移酶活性降低,DNA
存在异常的低甲基化。 4、神经精神疾病
精神分裂症和情绪障碍与DNMT基因相关。基因高甲 基化抑制脑组织中Reelin蛋白的表达,Reelin蛋白是 维持正常神经传递、大脑信息存储和突触可塑性所必 需的蛋白 。
组蛋白修饰及其功能(乙酰化,甲基化,磷酸化等)-于凯
当DNA与核小体尚未解开缠绕时,转录激活因子如糖皮质激素受体可以和DNA上相 应的反应元件(GRE)结合。当结合至GRE之后,糖皮质激素募集共激活因子如CBP到染 色体上的靶转录基因区。此时,共激活因子利用HAT活性使得结合在DNA启动子区域的核 心组蛋白乙酰化,进而使DNA与组蛋白结合减弱,核小体释放,转录因子和RNA聚合酶可 以与DNA上特异的启动子结合,启动靶基因的转录。
4
5
2. 组蛋白的甲基化
组蛋白甲基化是由组蛋白甲基化转移酶(histone methyl transferase,HMT)完成的。甲基化可发生在组 蛋白的赖氨酸和精氨酸残基上,而且赖氨酸残基能够 发生单、双、三甲基化,而精氨酸残基能够单、双甲 基化,这些不同程度的甲基化极大地增加了组蛋白修 饰和调节基因表达的复杂性。
9
10
组蛋白修饰的生物学意义
11
尤其是组蛋白乙酰化、甲基化修饰能为相关调控蛋白提供其在组蛋白上的附着位点,改变染色质结 构和活性。一般来说,组蛋白乙酰化能选择性的使某些染色质区域的结构从紧密变得松散,开放某 些基因的转录,增强其表达水平。而组蛋白甲基化既可抑制也可增强基因表达。乙酰化修饰和甲基 化修饰往往是相互排斥的。在细胞有丝分裂和凋亡过程中,磷酸化修饰能调控蛋白质复合体向染色 质集结。
通常,异染色质结构域组蛋白呈低乙酰化,常染色质结构 域组蛋白呈高乙酰化。
酵母组蛋白乙酰化与去乙酰化的调节
2
组蛋白乙酰化调节转录的机制
组蛋白乙酰化引起染色质结构改变及基因转录活性变化的至少包括以下几个方面: ①组蛋白尾部赖氨酸残基的乙酰化能够使组蛋白携带正电荷量减少,降低其与带负电荷的 DNA链的亲和性,导致局部 DNA与组蛋白八聚体解开缠绕,从而促使参与转录调控的各种 蛋白因子与DNA特异序列结合,进而发挥转录调控作用; ②组蛋白的N末端尾巴可与参与维持染色质高级结构的多种蛋白质相互作用,更加稳定了核 小体的结构。而组蛋白乙酰化却减弱了上述作用,阻碍了核小体装配成规则的高级结构(如 螺线管); ③组蛋白乙酰基转移酶(HAT)对相关转录因子或活化因子进行乙酰化修饰以调节基因的表 达。如 CBP/P300对P53的乙酰化可增强其特异性 DNA结合能力、转录激活能力,并延长其 半衰期。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
HAT识别底物的分子机制
r A. ‘hit-and-run’ model;
物 om r B. & C. attract-and生 .c hit’ models
做 oo r D. ‘targeted action’ 心 bi model
秀 r E. ‘relay’ model: 两个 物 - 专.b w HAT先后修饰底物
表观遗传学
第三章 组蛋白修饰
DNA Packing
r 1. 如何将10,000公里长的蚕
丝(半径~10-5米)装入一个篮 球中。
r 2. 蚕丝的体积:3.14*10-3m3
生物.com
r 3. 折叠、缠绕…
心做bioo
物秀- 专.b w
生 ww
染色体上不同的区域
r Euchromatin: 常染色质; r Heterochromatin: 异染色
5mC 物秀- 专.b w H4 乙酰化
H3 乙酰化
生 ww
组蛋白共价修饰的功能
r 基因转录、DNA损伤修复、DNA复制、染色体凝聚等
心做b生io物 生物w秀w- 专.b w
内容纲要
r 一、组蛋白的乙酰化
r r r r
r
六二 三 四 五、、 、 、 、组组 组 组 组蛋蛋 蛋 蛋 蛋白白白白白物密的的的的秀码甲磷泛SU-基酸素M专.化化化Ob心化w做b生io物
Gcn5/PCAF
Br, bromodomain;
物 om Nr, nuclear receptor生 .c interacting box; 做 oo CH, cysteine/histidine心 rich module; 秀 bi KIX, phospho-CREB 物 - 专.b w interacting module;
p300/CBP
Q, glutamine-rich
生 ww domain.
具有保守的HAT结构域
Bromodomain
r 1. An acetyl-lysine
binding domain
r 2. HATs修饰底物之后 可能与底物上的乙酰 化赖氨酸结合
心做b生io物
物秀- 专.b w
生 ww
心做b生io物
and H5-like domain;
秀- 专.b NEMM, N-terminal part of 物 w Enok, MOZ or MORF; 生 ww PHD, PHD zinc finger;
ED, glutamate/aspartate-
rich region;
SM, serine/methionine-rich
心做b生io物
物秀- 专.b w
核小体 生 ww
常染色质
异染色
组蛋白与核小体
心做b生io物 生物w秀w- 专.b w
Histone variants
心做b生io物 生物w秀w- 专.b w
组蛋白修饰
心做b生io物 生物w秀w- 专.b w
组蛋白修饰 (2)
生 ww
一、组蛋白的乙酰化
r 1. 通常发生在蛋白质的赖氨酸(K)上; r 2. 可逆的生化反应:
物 om | A. Histone acetyltransferase,HAT (>30) 生 .c | B. Histone deacetylase, HDAC (18) 心做bioo r 3. 分子效应:中和赖氨酸上的正电荷,增加组蛋 r 4白. 生与物DN学A功的物能排秀:斥-力专.b w 生 w | A. 基因转录活化 w | B. DNA损伤修复
酸化,促使GCN5修
秀- 专.b w 饰H3K14 物 E. SWI/SNF的BRG1 生 w 特异性识别H4K8; w TFIID的TAFII250识
别H3K9和H3K14, 从
而激活IFN-β
蛋白质乙酰化调控基因转录
r A. 乙酰化转录因子,使之与DNA结 合能力增强;
r B. 转录因子活化的结构域招募HATs
物 om 质 生 .c r E->H或H->称为染色质重塑
做 (Chromatin Remodeling) 心 bioo r 分子机理:DNA甲基化,
组蛋白修饰,染色质重塑复
物秀- 专.b w 合物的协同作用。 生 ww
常染色质与异染色质
r 1. 常染色质:基因表达
活跃的区域,染色体结 构较为疏松 r 2. 异染色质:基因表达 沉默的区域,染色体结 构致密
心做b生io物 生物w秀w- 专.b w
主要的功能基团
r Acetyl
r Methyl
心做b生io物
r Phosphory物l 秀- 专.b w
生 wwΒιβλιοθήκη r UbiquitinEpigenetic differences:monozygotic twins
生物.com
心做bioo
组蛋白的乙酰化
r 中和赖氨酸的正电荷,C=O具有一定的负电,能够增加 与DNA的斥力,使得DNA结构变得疏松,从而导致基因
生物.com 的转录活化 心做bioo
生物w秀w- 专.b w
HATs:转乙酰基酶
心做b生io物 生物w秀w- 专.b w
Gcn5/PCAF & p300/CBP
生 w r F. HIV Tat K50被p300 w 修饰后,招募PCAF,
再激活特定基因。 (Bromodomain)
人类IFN-β基因的激活
A. DNA code: 序列模
体、甲基化模式
B. GCN5结合到启动 子/增强子上
C. 修饰H4K8和H3K9 D. H3S10被RSK-2磷
心做b生io物
HATs: MYST(MOZ-Ybf2/Sas3-Sas2-TIP60)
Chromo, chromodomain;
具有保守的MYST结构域
Ser, serine-rich domain; CH, cysteine/histidine-rich motif;
H15, linker histones H1-
物 om 复合物; 生 .c r C. HAT复合物乙酰化组蛋白,打开
染色质;
心做bioo r D. 转录激活;
r E. HAT复合物中的共激活子也被乙
秀 酰化修饰; 物 - 专.b w r F. HAT复合物乙酰化之后离开,转
生 ww 录活性削弱。
组蛋白乙酰化:DNA repair
r 1. DNA损伤修复: