竹节参转录组使用密码子偏好性分析
豆科植物dxr基因密码子偏好性分析

Abstract 1 ̄deoxy ̄D ̄xylulose - 5 ̄phosphate reductoisomerase ( DXR) is one of the key enzymes in EMP pathwayꎬ involving in the bi ̄
osynthesis of terpenoids and anthraquinones. Due to the high content of terpenoids and ( or) anthraquinones in leguminous plantsꎬ it is
dxr 基因在酵母表达系统中具有高表达效率ꎬ仍需对其密码子进行优化ꎮ
关键词 豆科植物ꎻ1 ̄脱氧 ̄D ̄木酮糖 - 5 ̄磷酸还原异构酶ꎻ密码子偏好性ꎻ聚类分析
中图分类号 Q78ꎻQ943. 2
文献标识码 A
文章编号 2095 - 1736(2020)01 - 0030 - 05
Analysis of codon bias of dxr gene in leguminous plants
difference between yeast and E. coli was significantly lowerꎬ suggesting that yeast expression system is more suitable for the exogenous
expression system of dxr genes from leguminous plants. Howeverꎬ if the dxr genes of leguminous could be more efficiently expressed in
significant to study the codon bias of dxr gene in leguminous plants in order to understand the regulatory mechanism and further molecu ̄
兰科植物FNR基因的密码子偏好性分析

兰科植物FNR基因的密码子偏好性分析作者:李蓉谢析颖王雪晶苏立遥林玉玲郭容芳陈裕坤赖钟雄徐涵来源:《热带作物学报》2018年第06期摘要为揭示兰科植物FNR基因的特性和密码子偏好性,采用DNAMAN、CodonW和SPSS软件及 EMBOSS、SWISS-MODEL在线网站对17种兰科植物的FNR基因序列、氨基酸序列、同源区域及蛋白质三维结构进行分析。
结果表明:兰科植物FNR基因普遍具有在A/T (U)与G/C之间较弱的密码子偏好性;密码子的末位在A和T(U)之间存在显著的T (U)偏好性,在C和G之间存在显著的C偏好性;NADP结合域比非结合域有较小的密码子偏好性;基于CDS和氨基酸聚类结果比基于RSCU聚类更接近于植物的进化分类;自然选择的作用是导致兰科植物FNR基因的密码子使用偏好性的主要成因。
关键词兰科;FNR基因;密码子偏好性中图分类号 S682.31 文献标识码 ACodon Usage Bias of Ferredoxin-NADP+ Oxidoreductase (FNR)in OrchidaceaeLI Rong1, XIE Xiying1, WANG Xuejing1, SU Liyao1, LIN Yuling1, GUO Rongfang1,CHEN Yukun1, LAI Zhongxiong1*, XUHAN Xu1,2*1 Institute of Horticulture of Biotechnology, Fujian Agriculture and Forestry University,Fuzhou, Fujian 350002, China2 Institut de la Recherche Interdisciplinaire de Toulouse, Toulouse 313008, FranceAbstract For a good understanding of the codon usage bias of ferredoxin-NADP+ oxidoreductase (FNR) in Orchidaceae, the gene sequence, amino acid sequence, homologous region and three-dimensional protein structure of 17 FNR genes in Orchidaceae were analyzed by DNAMAN,CodonW, SPSS softwares, and EMBOSS, SWISS-MODEL online programs. The results showed that there was a low-level codon usage bias in FNR genes between A/T(U) and G/C;there existed a significant T(U) bias between A and T(U) and a significant C bias between C and G in the codons; NADP binding domain showed less codon bias than that of the non-binding domain; the cluster tree based on coding sequence (CDS) as well as amino acid sequences matched better with the plant evolution classification; the codon usage bias of FNR was mainly influenced by the natural selection in Orchidaceae.Key words orchidaceae; FNR gene; codon usage biasdoi 10.3969/j.issn.1000-2561.2018.06.015基因表达是细胞在生命活动过程中将储存在DNA双链中的遗传信息转录、翻译为具有生物活性的蛋白质的过程。
密码子偏好性分析

摘 要 脂多糖结合蛋白(lipopolysaccharide-binding protein, LBP)是机体识别革兰氏阴性菌内毒素并启 动免疫反应的关键因子。为了了解 LBP 基因的密码子使用特性,为其选择合适的受体动物以及最佳外源 表达系统提供依据,本研究运用 CHIPS、CUSP 和 CodonW 在线程序分析自主电子克隆的猪(Sus scrofa) LBP 基因(GenBank 登录号: NM-001128435.1)的密码子偏好性,并与猪 8 种抗病相关基因、模式生物基因 组以及其他物种 LBP 基因相比较。结果表明,猪 LBP 基因大部分偏好使用以 G/C 结尾的密码子,27 种偏 好密码子(相对使用度(RSCU)>1)中偏好性较强的有 GCC、CAC、CTG 和 TCC(RSCU≥2),而猪 8 种抗病 相关基因有 23 种偏好密码子,全部以 G/C 结尾,并且偏好性较强的密码子有 GCC、ATC、CTG 和 GTG;通 过比较 14 种动物的 LBP 基因密码子偏好性,发现 14 个物种的 LBP 基因表达水平一般,并且都偏好以 G/C 结尾的密码子;聚类分析发现,偶蹄目猪与 2 种食肉目动物(猫(Felis catus)和狗(Canis))聚为一类,与系统 分类关系不一致;在密码子的使用频率上,猪 LBP 基因与小鼠(Mus musculus)基因组的差异小于大肠杆菌 (Escherichia coli)和酵母菌(Saccharomyces)等 2 种模式生物基因组,故小鼠更适合作为该 LBP 基因的外源表 达宿主。本研究结果为 LBP 基因在动物遗传改良中选择合适的受体动物、选择最佳的外源表达系统以及 提高其表达水平提供一定的理论依据。 关键词 猪,脂多糖结合蛋白基因(LBP),密码子偏好性
南欧大戟叶绿体基因组密码子偏好性分析

櫄櫄櫄櫄櫄櫄櫄櫄櫄櫄櫄櫄櫄櫄櫄櫄櫄櫄櫄櫄櫄櫄櫄櫄櫄櫄櫄櫄櫄櫄櫄櫄櫄櫄櫄櫄櫄櫄櫄櫄櫄櫄櫄櫄櫄櫄 济南:山东大学,2007.[16]王晓东.转基因小麦抗旱性生理生化及农艺性状鉴定[D].杨凌:西北农林科技大学,2016:7-48.[17]闫建俊,白云凤,左静静,等.转基因马铃薯外源基因插入位点分析及检测方法的建立[J].分子植物育种,2020,18(16):5361-5366. [18]游 朝,晁朝霞,姚正培,等.转MvNHX1和MvP5CS基因棉花耐盐抗旱性比较与育种价值分析[J].棉花学报,2015,27(3):198-207.[19]易小平,谭燕华,彭存智,等.转基因作物安全评价的检测技术[J].热带生物学报,2015,6(1):98-104.[20]康 丹,方小艳,游腾飞,等.染色体步移技术克隆已知序列侧翼启动子的研究进展[J].农业生物技术学报,2013,21(3):355-366. [21]赵才美,黄兴奇,殷富有,等.水稻NAC转录因子家族的研究进展[J].植物科学学报,2020,38(2):278-287.[22]段晓亮,许兰杰,刘志勇,等.转基因小麦外源基因插入位点初步分析及检测方法的建立[J].粮油食品科技,2014,22(4):76-81.[23]姜子焱.梭梭HaNAC38、HaNAC42启动子克隆和转录因子特性分析[D].乌鲁木齐:新疆农业大学,2018:3-37.[24]涂松林,施爱民.我国转基因棉花研究与应用进展[J].江西棉花,2001,23(1):9-13.[25]陈秀兰,张玉忠,张 军.棉花分子育种研究进展[J].棉花学报,1997,9(1):5-8.[26]王瑞芳,胡银松,高文蕊,等.植物NAC转录因子家族在抗逆响应中的功能[J].植物生理学报,2014,50(10):1494-1500.[27]王志霞.转基因棉花研究进展[J].江苏农业学报,2003,19(2):74.[28]林 清,彭于发,吴 红,等.转基因作物及产品检测技术研究进展[J].西南农业学报,2009,22(2):513-517.[29]王淑君,曲延英,倪志勇,等.转CarNAC1基因可提高棉花的抗旱性[J].干旱地区农业研究,2018,36(4):272-281.余 涛,蒲 芬,管 芹,等.南欧大戟叶绿体基因组密码子偏好性分析[J].江苏农业科学,2023,51(15):35-41.doi:10.15889/j.issn.1002-1302.2023.15.006南欧大戟叶绿体基因组密码子偏好性分析余 涛,蒲 芬,管 芹,范 敏(大理大学药学院,云南大理671000) 摘要:为了揭示南欧大戟叶绿体基因组密码子使用偏性及形成原因,以南欧大戟叶绿体基因组中长度大于300bp的非重复蛋白质编码序列为研究对象,利用CodonW1.4.1、CUSP在线程序等生物信息学分析工具对其密码子偏性及使用模式进行分析。
大豆基因组和转录组的核基因密码子使用偏好性分析

1
1.1
材料与方法
数据的获取 基因组水平的基因数据是从大豆基因组数据库 Phytozome (/soybean) 批 量 下载 (batch download)的大豆全基因组的高置信蛋白 编码基因序列 , 以及叶片和种子的特异表达基因序 列。根据大豆基因组数据库对已知大豆基因功能的 注释和未知基因通过同源比对等方法的功能预测 ,
1.2
表 1 叶片特异表达基因信息 Table 1 Information of leaf-specific genes 基因名称 Gene name Glyma02g29170 Glyma09g17060 Glyma10g38910 Glyma20g28890 Glyma10g38910 Glyma20g28890 Glyma04g04450 Glyma14g08560 Glyma17g36530 Glyma05g01770 Glyma11g09920 Glyma12g02260 染色体 Chr. 2 9 10 20 10 20 4 14 17 5 11 12 功能注释 Function annotation Protein phosphatase 2C Protein phosphatase 2C Chlorophyll A-B binding protein Chlorophyll A-B binding protein Chlorophyll A-B binding protein Chlorophyll A-B binding protein Protein of unknown function (DUF3223) CYTOCHROME P450 Glyma17g36530.1 Aldehyde dehydrogenase family Domain of unknown function (DUF3411) Domain of unknown function (DUF3411)
不同植物FERONIA基因密码子使用偏好性分析
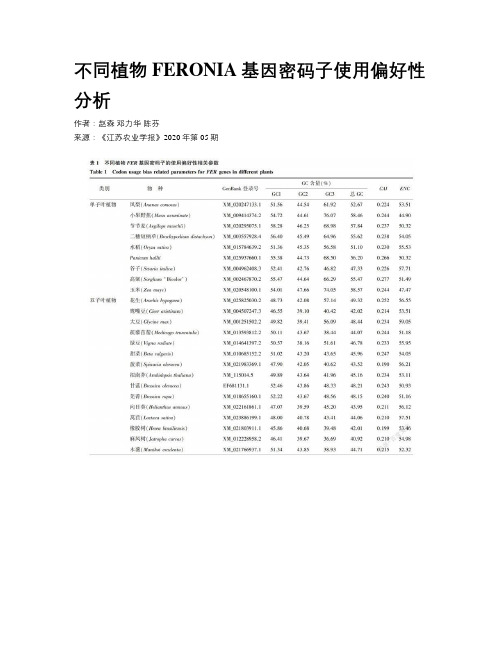
不同植物FERONIA基因密码子使用偏好性分析作者:赵森邓力华陈芬来源:《江苏农业学报》2020年第05期摘要:为了了解植物FERONIA(FER)基因在长期进化过程中所形成的密码子使用模式及它们之间的亲缘关系,采用Codon W、CUSP在线程序和SPSS等分析软件对不同植物的FER基因密码子进行碱基组成分析、对应性分析、PR2-Plot分析、中性绘图分析和ENC-Plot 分析并进行聚类分析。
结果表明,单子叶植物FER基因密码子的平均G+C含量高于双子叶植物,且单子叶植物偏好使用以G/C结尾的密码子,而双子叶植物偏好使用以A/T结尾的密码子。
对应性分析结果显示,在单子叶植物、双子叶植物中,向量轴1(Axis 1)可解释的变异分别占总变异的42.25%、28.56%。
在单子叶植物中,Axis 1与总G+C含量、GC3含量、G3含量、C3含量呈极显著正相关(P<0.01),而在双子叶植物中则呈极显著负相关(P<0.01)。
不同的碱基组成是单子叶植物和双子叶植物形成密码子使用偏好性的重要原因。
PR2-Plot分析结果、中性绘图结果和ENC-Plot分析结果表明,自然选择是形成植物FER基因密码子使用偏好性的主要驱动力。
基于编码序列(Coding sequence,CDS)构建的系统进化树比基于同义密码子相对使用度(RSCU)得出的聚类分析结果更接近传统的分类结果。
本研究结果可为进一步开展植物分子进化和基因工程研究奠定基础。
关键词:FERONIA基因;密码子使用偏好性;分子进化中图分类号:Q754;Q755文献标识码:A文章编号:1000-4440(2020)05-1073-09Abstract:To understand the usage pattern of codons formed in the long-term evolutionary process of FERONIA (FER) genes in plants and their genetic relationships, base composition analysis, correspondence analysis, PR2-Plot analysis, neutral plotting analysis and ENC-Plot analysis on FER gene codons of different plants were conducted by using Codon W, on line program of CUSP and analysis software like SPSS, and the cluster analysis was conducted. The results showed that the average content of G and C in FER gene codons of monocots was higher than that of dicots, and the monocots preferred G/C ending codons while dicots preferred A/T ending codons. The results of correspondence analysis showed that, Axis 1 accounted for 42.25% and 28.56% of the total variation in monocots and dicots respectively. Axis 1 presented significantly positive correlation (P<0.01) with contents of total G+C, GC3, G3 and C3 in monocots,while in dicots the correlation between Axis 1 and contents of total G+C, GC3, G3, C3 was significantly negative respectively. Different nucleotide composition is the important reason in the formation of codon usage preference in monocots and dicots. Results of PR2-Plot analysis, neutral plotting analysis as well as ENC-Plot analysis showed that, natural selection was the main driving force in the forming of codon usage preference of FER genes in plants. Phylogenetic tree constructed by coding sequence (CDS) was more similar with traditional classification results than the phylogenetic tree constructed by relative synonymous codon usage (RSCU). The results can lay foundation for further research on molecular evolution and gene engineering.Key words:FERONIA gene;codon usage preference;molecular evolution根據中心法则,遗传信息流的方向是从DNA经RNA到蛋白质,密码子则是生物体内核酸信息和蛋白质信息相互沟通的桥梁。
不同哺乳动物密码子偏性及聚类分析
不同哺乳动物密码子偏性及聚类分析在雌性哺乳动物整个生殖周期过程中,卵母细胞只能来源于原始卵泡库[1]。
有研究表明,原始卵泡形成过程受激素、转录因子和相关通路的介导[2]。
生长分化因子9(growth differentiation factor 9,GDF9)作为转化生长因子β超家族成员之一,在卵泡的发育过程中有关键的调节作用[3]。
密码子是DNA或RNA的碱基序列与其编码蛋白序列之间的对应关系[4]。
编码相同氨基酸的密码子为同义密码子,在蛋白合成过程中,同义密码子的使用频率存在差异,且物种和基因对某一种或几种密码子的使用具有偏好性[5]。
对基因密码子偏好参数进行分析,能更好地理解和研究基因水平转移和基因家族分化的发生[6]。
因此对密码子偏好性的研究可在分子的角度为GDF9基因序列特征、分类和遗传进化规律提供重要信息。
前人相关研究表明,GDF9基因具有刺激颗粒细胞减数分裂、抑制素的生成作用[7],并影响卵泡发育和生殖功能[8]。
Wang等发现通过siRNA敲除GDF9后能抑制仓鼠原始卵泡的形成,但添加GDF9纯品培养卵巢会加速原始卵泡的形成[9]。
在人、啮齿类、牛、绵羊和有袋类动物卵巢卵母细胞中,GDF9基因特异性表达,但该基因在山羊卵母细胞和黄体中可同时表达[10-11]。
GDF9基因还能通过多个信号通路促进颗粒细胞增殖过程[12]。
马会明等通过RNA干扰使该基因表达沉默,也能抑制颗粒细胞增殖过程[13]。
这些研究通过探讨不同哺乳动物已被克隆的GDF9基因,并在卵巢卵母细胞中进行了相关表达分析,但未开展密码子使用偏性的研究,这不利于其异源表达和遗传转化等后续试验的进行。
GDF9基因密码子使用偏好性的研究能为该基因的分类和进化提供重要信息。
本研究利用CodonW软件分析不同哺乳?游?GDF9基因对密码子的使用情况,基于GDF9基因最小进化法和同义密码子相对使用度的欧式平方距离系数建立聚类关系,为GDF9基因功能的深入研究提供参考依据。
黄荆坪竹根椒叶绿体基因组密码子使用偏好性及影响因素分析
龚意辉,唐诗眙,周桂花,等.黄荆坪竹根椒叶绿体基因组密码子使用偏好性及影响因素分析[J].江苏农业科学,2023,51(20):28-34.doi:10.15889/j.issn.1002-1302.2023.20.005黄荆坪竹根椒叶绿体基因组密码子使用偏好性及影响因素分析龚意辉1,唐诗眙1,周桂花1,陈 磊1,张 斌1,李 鹏2(1.湖南人文科技学院农业与生物技术学院,湖南娄底417000;2.湘潭市农业科学研究所,湖南湘潭411134) 摘要:为了解黄荆坪竹根椒(CapsicumannuumL.cvHuangJingping)叶绿体基因组密码子使用模式,以其51条蛋白编码序列(codingsequence,CDS)为研究对象,采用CodonW1.4.2、CUSP、Excel2010、SPSS16.0等软件对黄荆坪竹根椒密码子的GC含量、有效密码子值(ENC)、RSCU、中性绘图、ENC-plot、PR2-plot、最优密码子及其影响因素进行详细的分析。
结果表明,黄荆坪竹根椒密码子第1位(GC1)、第2位(GC2)、第3位(GC3)中的GC含量分别为42 68%、36.25%、27.89%,其中GC1>GC2>GC3,表明密码子第3位碱基以A/U结尾为主;ENC值介于34.10%~54 31%之间,平均值为46.55%,表明黄荆坪竹根椒密码子偏好性较弱。
相关性分析结果表明:(1)GC1与GC2呈极显著正相关,GCall与GC1、GC2、GC3均为极显著正相关;(2)ENC与GC1、GC2均不存在显著相关关系,但与GC3存在极显著相关关系。
RSCU分析结果显示:在黄荆坪竹根椒叶绿体基因组中共有30个高频密码子的RSCU>1,其中以U、A、G结尾的高频密码子数分别为16、13、1个,表明黄荆坪竹根椒密码子第3位偏好以A或U结尾。
ENC-plot绘图分析结果表明,黄荆坪竹根椒基因组中大多数基因位于标准曲线下方,有13个基因的ENC比值介于-0.05~0.05之间,说明选择压力主要影响黄荆坪竹根椒密码子的偏好性;PR2-plot分析结果表明,黄荆坪竹根椒叶绿体基因组中大部分基因位于平面图的右下方,即T>A、G>C,表明碱基突变、自然选择压力等多种因素共同影响黄荆坪竹根椒密码子的偏好性。
密码子偏好表
密码子偏好表是指生物体内不同基因在不同组织或不同细胞类型中的密码子使用偏好性。
它反映了基因在不同环境下的表达和翻译效率。
密码子偏好性对于理解基因表达调控、药物设计和生物信息学等领域具有重要意义。
首先,从遗传密码的特性来看,密码子偏好性是必然存在的。
生物体使用的密码子有3个,分别代表不同的氨基酸。
由于自然界中氨基酸的种类有限,因此不同的密码子在生物体内出现的频率必然有所不同。
这反映了基因在不同环境下的表达和翻译效率。
其次,不同基因在不同组织或不同细胞类型中的密码子偏好性存在差异。
这种差异可能是由于基因在特定组织或细胞类型中的表达调控机制所致。
例如,某些基因在某些组织中的表达可能受到转录因子或RNA加工机制的影响,而这些机制可能会影响密码子的使用偏好性。
再者,密码子偏好性受到生物体内各种因素的影响,如代谢状态、环境压力、营养条件等。
这些因素可能通过影响基因的表达和翻译效率来影响密码子的使用偏好性。
此外,遗传变异和进化也可能会影响密码子的使用偏好性,从而使不同的物种和种群表现出不同的密码子偏好性。
具体来说,某些密码子在生物体内可能更常见于编码某些特定的氨基酸,而其他密码子则可能较少出现。
这可能是由于某些氨基酸在生物体内的需求较高,因此在基因表达和翻译过程中更频繁地使用某些密码子。
此外,某些氨基酸可能与特定的辅因子或信号分子结合,从而影响其翻译效率,这也可能导致某些密码子的偏好性。
总之,密码子偏好表反映了生物体内不同基因在不同组织或不同细胞类型中的密码子使用偏好性。
这种偏好性是基因在不同环境下的表达和翻译效率的体现,同时也受到各种因素的影响。
密码子偏好性对于理解基因表达调控、药物设计和生物信息学等领域具有重要意义。
通过研究密码子偏好性,我们可以更好地了解生物体的基因表达机制和蛋白质合成过程,从而为药物设计和疾病治疗提供新的思路和方法。
密码子偏好性的相关研究及方法分析
密码子偏好性的相关研究及方法分析
冉然;张晓明;宛涛;伊卫东
【期刊名称】《草原与草业》
【年(卷),期】2022(34)3
【摘要】密码子偏好性指的是物种对编码相同氨基酸的同义密码子的不均等使用。
这样的现象和生物功能分子蛋白质和遗传信息载体分子DNA有相关联系,因此对
其的研究具有非常重要的意义。
本文概述了密码子偏好性的研究进展,实现密码子
偏好性的生物学基础,并简要介绍了研究密码子偏好性的意义以及研究分析方法以
及相关研究在草业科学中应用的意义。
【总页数】7页(P5-10)
【作者】冉然;张晓明;宛涛;伊卫东
【作者单位】内蒙古农业大学草原与资源环境学院/草地资源教育部重点实验室/农业农村部饲草加工高效利用重点实验室/内蒙古自治区草地管理与利用重点实验室【正文语种】中文
【中图分类】S812;Q75
【相关文献】
1.基于相关分析的超偏载装置精度评价方法研究
2.密码子偏性的分析方法及相关研究进展
3.密码子偏性分析方法及茶树中密码子偏性研究进展
4.应用脑电信号偏直
线相关分析方法研究颞叶内侧癫痫网络连接机制5.应用脑电信号偏直线相关分析
方法研究颞叶内侧癫痫网络连接机制
因版权原因,仅展示原文概要,查看原文内容请购买。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
梁 娥ꎬ齐敏杰ꎬ丁延庆ꎬ等.竹节参转录组使用密码子偏好性分析[J].江苏农业科学ꎬ2019ꎬ47(2):59-63.doi:10.15889/j.issn.1002-1302.2019.02.013
竹节参转录组使用密码子偏好性分析
梁 娥1ꎬ齐敏杰1ꎬ丁延庆2ꎬ张 来2(1.贵州师范大学生命科学学院ꎬ贵州贵阳550000ꎻ2.安顺学院农学院ꎬ贵州安顺561000)
摘要:竹节参是我国珍稀濒危中药材ꎬ研究其基因密码子使用模式ꎬ可为利用基因工程技术实现人参皂苷异源生物合成及竹节参分子育种改良提供理论依据ꎮ以竹节参转录组测序结果为数据来源ꎬ筛选编码蛋白基因序列(codingsequenceꎬ简称CDS)碱基数不小于300bp的11199条完整开放阅读框序列作为研究对象ꎬ利用Codon和SPSS软件分
别统计竹节参基因密码子GC含量、密码子第3位的(C+G)含量(GC3)和密码子第1、第2位(G+C)含量的平均值(GC12)、同义密码子的相对使用度(RSCU)、有效密码子数(ENC)等密码子偏好性指标ꎬ通过中性绘图(GC12vs.
GC3)、PR2绘图和ENC-GC3s绘图分析影响竹节参密码子使用模式的因素ꎮ结果表明ꎬ竹节参基因的平均GC、GC12
和GC3s含量分别为44.67%、46.97%和39.80%ꎬ其密码子使用模式受到突变和选择等多重因素的影响ꎬ确定了31个竹节参最优密码子ꎬ除了UUG外ꎬ其余最优密码子均以A或T结尾ꎮ竹节参密码子使用模式与大肠杆菌和酿酒酵母相比差异较大ꎬ选取毕赤酵母作为竹节参基因的异源表达宿主更为合适ꎮ 关键词:竹节参ꎻ转录组ꎻ密码子使用模式ꎻ最优密码子
中图分类号:Q755ꎻS567.5+10.1 文献标志码:A 文章编号:1002-1302(2019)02-0059-05
收稿日期:2017-08-27基金项目:国家自然科学基金(编号:31660252)ꎻ贵州省优秀青年科技人才专项(编号:黔科合人字[2015]18号)ꎻ贵州省教育厅创新群体重大研究项目(编号:黔教合KY[2016]049号)ꎮ作者简介:梁 娥(1994—)ꎬ女ꎬ贵州遵义人ꎬ硕士研究生ꎬ主要从事微生物学研究ꎮE-mail:1013653671@qq.comꎮ通信作者:张 来ꎬ博士ꎬ教授ꎬ主要从事植物次生代谢调控研究ꎮE-mail:975575681@qq.comꎮ
遗传密码子是生物体DNA与蛋白质之间信息传递的基本单位ꎬ具有简并性ꎬ即同一氨基酸有多个对应的密码子ꎬ编码同一种氨基酸的密码子叫作同义密码子ꎬ同义密码子在同一物种不同基因间或不同物种内的使用频率大有不同ꎬ这种不均衡使用模式称为密码子使用偏好性ꎬ通常把使用频率较高的一种或几种同义密码子称为最优密码子[1-2]ꎮ研究显示ꎬ不同物种之间基因密码子偏好性是由突变压力(如GC含量、基因碱基组成)和自然选择作用(如翻译起始信号、基因表达水平、蛋白结构与长度、tRNA丰度等)引起的ꎬmRNA的二级结构及其稳定性、翻译的速度和准确度、蛋白质折叠等因素也与密码子的偏好性有关[3-4]ꎮ对物种密码子偏好性开展研究ꎬ有助于理解物种进化发展及密码子使用偏好性的调控机制ꎮ密码子偏好性在基因异源表达研究方面也显示了重要作用ꎬ基因的表达量越大ꎬ其密码子偏好性越强[5]ꎬ根据这一原理ꎬ替换基因低表达密码子可以提高外源基因表达量ꎬ同时根据密码子使用偏好性可以选择更为合适的宿主表达系统ꎮ有报道表明ꎬ可通过优化密码子的方式来提高外源基因在宿主细胞中的表达量[6-7]ꎮ周宗梁等通过优化密码子的方法提高了苏云金芽孢杆菌基因cry1Ah在玉米和水稻中的表达量[8-9]ꎻ杨金玲等通过优化蝎毒镇痛活性肽基因BmKAngM1ꎬ将其导入毕赤酵母后该基因表达量得到显著提高[10]ꎻ通过使用最优密码子ꎬ在草菇[11]、拟南芥[12]、川母贝[13]、菠萝[14]等生物中均得到了很好的研究成果ꎮ竹节参(PanaxjaponicusC.A.Mey)为多年生草本植物ꎬ属于五加科(Paeoniaceae)人参属ꎬ是我国珍稀濒危的“七类中草药”之一ꎬ具有抗炎、延缓衰老、降血糖等药理作用ꎬ有着极高的药用和保健价值ꎮ竹节参中富含活性物质三萜皂苷ꎬ也是其特征性成分ꎮ目前在竹节参种质资源[15-16]、毛状根的
培养[17]、基因工程代谢的调控[18]、生药学鉴定[19]、精油成分
分析[20]和三萜皂苷代谢追踪[21]等几个研究领域已经开展了许多研究工作ꎬ但直接从竹节参中通过分离提取的方法获得三萜皂苷对资源消耗极大ꎬ技术难度较高ꎮ若利用生物合成的方法则可以很好地解决这一困难ꎬ实现有效成分的生物合成ꎬ选择适合关键酶基因高效表达的异源表达系统是一个重要步骤ꎮ本研究以竹节参转录组数据为材料ꎬ通过分析竹节参基因密码子组成的各项指标ꎬ研究竹节参表达基因密码子使用偏好性及其影响因素ꎬ以期为竹节参相关基因表达系统的选择及分子育种提供理论基础ꎮ
1 材料与方法1.1 数据来源竹节参转录组数据来源于文献[22]ꎬ通过Perl语言程序对竹节参转录组数据进行过滤筛选ꎬ筛选出碱基数≥300bp的蛋白质编码序列共11199条ꎬ作为密码子分析的数据来源ꎮ本研究中使用到的大肠杆菌(Escherichiacoli)、酿酒酵母(Saccharomycescerevisiae)和毕赤酵母(Pichiapastoris)的密码
子偏好性数据来自CodonUsagedatabase(http://www.kazusa.or.jp/codon/)ꎮ
1.2 竹节参基因GC含量分析及中性绘图
利用CodonW1.4.2(http://codonw.sourceforge.net/)统计分析竹节参基因密码子的碱基组成规律ꎬ测得鸟嘌呤和胞嘧啶总体含量G+C、密码子第3位碱基组成(A3、G3、C3、
—95—江苏农业科学 2019年第47卷第2期T3)、密码子第3位的C+G数(GC3)和密码子第1、第2位的平均G+C含量(GC12)等数据ꎮ本研究以密码子第1、第2个碱基的G+C平均含量为纵坐标ꎬ以密码子第3个碱基的G+C含量为横坐标ꎬ通过中性绘图探究影响密码子偏好性的因素ꎮ分析密码子第1、第2位与第3位碱基组成的相关性ꎬ来分析自然选择与压力突变对基因密码子使用模式的影响[23]ꎮ1.3 竹节参基因有效密码子数分析参照Fuglsang的方法[24]ꎬ利用有效密码子数来衡量竹节参基因单个密码子使用的偏好程度ꎮ基因有效密码子数(effectivenumbercodonꎬ简称ENC)是衡量基因密码子偏好性的一个重要指标ꎬ数值范围为20(每个氨基酸只使用1个同义密码子的极端偏好情况)到61(每个同义密码子被平均使用的无偏好情况)ꎮ研究表明ꎬ当ENC值≤35时ꎬ基因密码子的使用偏好性越强[25]ꎮ同时以ENC值为纵坐标ꎬGC3s为横坐标进行ENC绘图ꎬ分析各基因密码子的使用特征ꎬ研究竹节参密码子偏好性和基因碱基组成之间的关系ꎮ1.4 同义密码子相对使用度及最优密码子分析参照Sharp等的方法[26]ꎬ采用同义密码子的相对使用度(RSCU)作为衡量竹节参密码子使用偏好性的指标ꎮ使用3个指标表示:密码子使用无偏好性(RSCU=1)ꎻ密码子使用度较大(RSCU>1)ꎻ密码子使用频率较低(RSCU<1)ꎮ利用高表达优越密码子分析方法[27]ꎬ统计所有基因的ENC值、有序数据集的上下10%区间形成高RSCU集合和低RSCU集合ꎬ进行最优密码子分析ꎮ根据2个子集的ΔRSCU值及卡方检验确定最优密码子ꎮ1.5 PR2绘图分析采用PR2绘图进行密码子奇偶偏好分析[23]ꎬ分别统计竹节参基因中A3/(A3+T3)与G3/(G3+C3)数据ꎬ以此为纵坐标和横坐标ꎬ通过平面图显示各基因碱基的组成ꎮ为使编码密码子第3位碱基对A/T或T/A和G/C或C/G的突变不均衡ꎬPR2绘图时只选择亮氨酸(Leuꎬ密码子:CTA、CTC、CTG、CTT)、脯氨酸(Pro)、精氨酸(Argꎬ密码子:CGA、CGC、CGG、CGT)、丝氨酸(Serꎬ密码子:TCA、TCC、TCG、TCT)、丙氨酸(Ala)、缬氨酸(Val)、苏氨酸(Thr)和甘氨酸(Gly)等遗传密码子进行分析ꎮ2 结果与分析2.1 GC含量分析以及中性绘图竹节参转录组数据中共有11199条完整的开放阅读框序列ꎬ利用CodonW对完整的开放阅读框序列进行密码子使用模式分析ꎮ结果(表1)表明ꎬ所有完整的开放阅读框序列总长度为12006732bpꎬN50=1332bpꎬ平均GC含量为44.67%ꎬGC含量范围为30.3%~63.8%ꎮ竹节参基因组平均GC含量低于大肠杆菌基因组平均GC含量(52.35%)ꎬ高于酿酒酵母基因组平均GC含量(39.77%)和毕赤酵母基因组平均GC含量(42.73%)ꎮ竹节参基因组密码子第1、第2位碱基GC含量的变化范围为31.7%~71.8%ꎬ平均GC含量为4697%ꎮ密码子的第3位上碱基GC含量的变化范围为107%~83.9%ꎬ平均GC含量是39.80%ꎮ密码子第3位碱基上A、T的使用频率分别是33.09%、41.27%ꎬ略高于C、G的使用频率(分别为23.61%、27.57%)ꎬ说明竹节参基因密码子对以A和T结尾的密码子的使用偏好程度较大ꎬ而对以G和C结尾的密码子的使用偏好程度较小ꎮ密码子第3位碱
基上的平均GC含量(39.80%)略低于毕赤酵母基因密码子第3位碱基的平均GC含量(42.16%)ꎬ偏高于酿酒酵母密码子第3位碱基的平均GC含量(38.10%)ꎬ大大低于大肠杆菌遗传密码子第3位碱基的GC平均含量(55.62%)ꎮ研究表明ꎬ竹节参对密码子使用并无特殊碱基偏好ꎬ其密码子使用模式与大肠杆菌和酿酒酵母差异较大ꎬ与毕赤酵母差异略小ꎮ如图1所示ꎬ中性绘图分析表明ꎬ竹节参密码子GC12取值范围为31.7%~71.8%ꎬGC3的取值范围为10.7%~83.9%ꎬGC3s与GC12的相关系数为0.062ꎬ回归系数为0.48ꎬ表明并
无显著相关性ꎬ说明竹节参基因密码子使用模式主要受到自然选择的影响ꎮ
表1 不同物种密码子平均GC含量转录核酸中的GC密码子
平均GC含量(%)
竹节参酿酒酵母毕赤酵母大肠杆菌平均含量44.6739.7742.7352.35GC339.8038.1042.1655.62
GC1246.9740.6143.0250.72
2.2 竹节参基因有效密码子数分析研究显示ꎬ竹节参ENC值一般在25.88~61.00ꎬENC平均值为53.04ꎮ通常将ENC值=35作为衡量密码子偏好性强弱的标准[22]ꎮ竹节参基因有13条ENC值小于35ꎬ表明竹
节参基因整体水平密码子偏好性较低ꎬ只有少数基因具有密码子偏好性ꎮ由表2的ENC相关性分析可知ꎬGC与GC12、GC3之间的相关性均达极显著水平ꎬGC12和GC3之间并没