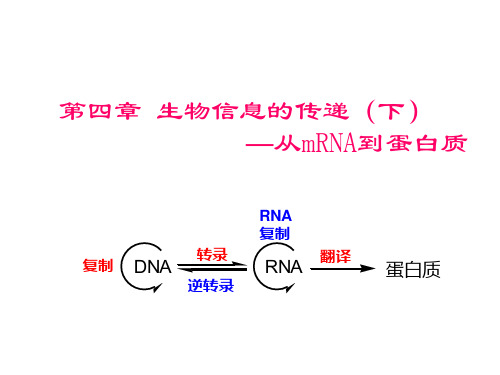
大学分子生物学经典课件第四章 生物信息的传递(下) 从mRNA到蛋白质
合集下载
第四章生物信息的传递(下)1

起始密 码子
2020/9/19
4、密码的摆动性
1966年,Crick提出摆动假说(Wobble hypothesis) •tRNA上的反密码子与mRNA上的密码子配对 时,密码子的第一位、第二位碱基配对是严格 的,第三位碱基可以有一定变动,这种现象称 为密码的摆动性或变偶性(wobble)。 •I(肌苷,次黄嘌呤核苷)A、U、C配对。
密码,直接测出三联体对应的氨基酸; (5)到1966年,遗传密码全部破译。
2020/9/19
遗传密码的破译
1.制备E.coli无细胞合成体系,以均聚物、随机共聚 物和特定序列的共聚物模板指导多肽的合成。 •多聚同一核苷酸的翻译 •多聚重复核苷酸的翻译
2.核糖体结合技术
2020/9/19
核糖体结合技术
tRNA:运送特定氨基酸到核糖体上合成 蛋白质。
rRNA:核糖体的组成元件。
2020/9/19
Protein synthesis uses three types of RNA
3` 5`
2020/9/19
mRNA
2020/9/19
原核生物mRNA与真核生物mRNA结构比较
Prokaryotic mRNA 核糖体可以不从mRNA上解离连续合成三个蛋白质
(二)同工tRNA
携带相同氨基酸而反密码子不同的 一组tRNA称为同功受体tRNA
原因:tRNA 的数目(>20余种)大于氨基酸数目
2020/9/19
同工tRNA的特性:
同工tRNA既要有不同的反密码子以识别该氨基 酸的各种同义密码,又要有某种结构上的共同 性,能被AA- tRNA合成酶识别.
2020/9/19
2020/9/19
大肠杆菌无义突变的校正tRNA
04郭江波分子生物学讲义-第四章
错义抑制
错义突变
AUG AGA UAA AUG
错义突变
GGA UAA AUG AGA UAA
UCU
CCU 抑制突变
UCU
Arg
Gly
Gly
图 14-18 反密码子发生突变可抑制错义突变
异亮氨酸(Ile)和缬氨酸 (Val)
异亮氨酸--tRNA合成酶对异亮氨酸的亲和力比缬
氨酸大225倍,体内缬氨酸浓度比异亮氨酸高5
nucleotide present in the third codon
position.
2、通用性与特殊性
• 蛋白质生物合成的整套密码,从原核生物到 人类都通用。 • 已发现少数例外,如动物细胞的线粒体、植 物细胞的叶绿体。
3、 连续性
• 三个核苷酸编码一个氨基酸。 • 三联子密码是非重叠(non-overlapping)和连
第四章 生物信息的传递(下) —从mRNA到蛋白质
RNA 复制 复制
DNA
转录 逆转录
RNA
翻译
蛋白质
• 蛋白质是生物信息通路上的终产物,一个 活细胞在任何发育阶段都需要数千种不同 的蛋白质。
• 活细胞内时刻进行着各种蛋白质的合成、 修饰、运转和降解反应。 • 蛋白质由20中氨基酸组成。
翻译:指将mRNA链上的核甘酸从一个特定的起始位
• Arg, Ser, Leu 6 codons • Gly, Thr, Ala, Val, Pro 4 codons • Met, Trp 1 codon
Third base degeneracy
describes the lesser
e
点开始,按每三个核苷酸代表一个氨基酸的原则, 依次合成一条多肽链的过程。 蛋白质合成的场所是 蛋白质合成的模板是 核糖体 mRNA tRNA
生物信息的传递--从RNA到蛋白质

(2) P位点:肽酰基-tRNA位点
❖大部分在小亚基内 ,小部分在大亚基内 16SrRNA的3‘末端 ❖L2、L27及L14 、L18、L24 、L33 ❖能够与起始tRNA(Pro中 fMet-tRNAf)相结合
(3)A位点:氨酰基-tRNA位点
❖在大亚基上 ❖ A位点内mRNA表面只对特定的aa—tRNA分子表 现出特异性。
G → C/U (Py) U*(经过修饰) → G/A(Pu)
起 始
Ile
Arg
End of section 1
第二节 基本元件
一、核糖体及rRNA的结构
❖翻译场所,含大小亚基; ❖Prok中转录与翻译偶联, 多聚核糖体; ❖Euk中多与细胞骨架和内 质网膜结合。
图示尺寸比较显示核糖 体足以结合tRNA和mRNA
1、 核糖体的结构
有注
2、核糖体的装配
3、核糖体的活性位点
❖根据功能将核糖体上的活性部位分为两类
➢翻译区域 7个活性位点 占2/3 ➢逐出位点 2个位点(多肽的逐出)占1/3
膜
E2 位点
(1)mRNA结合位点
❖位于小亚基的头部; ❖S1 (防止mRNA链内碱基对的形成) ❖16S rRNA 3’端
主要内容
第一节 遗传密码 第二节 基本元件 第三节 肽链的合成 第四节 蛋白质前体的加工与转运机制
❖基因表达的第二步;
前言
❖tRNA,rRNA和mRNA共同参与;
❖tRNA起“译员”的作用;
❖tRNA既是密码子的受体,也是氨基酸的受体;
❖tRNA通过其自身的反密码子识别密码子;
❖(三联体)密码子有自身的特性 ➢连续性(不重叠性,无标点性)
❖Crick 1961 提出, 又称 “变偶假说”
从mRNA到蛋白质生物信息的传递讲课文档

稀有碱基出现的频率高,对于维持反密码环的稳定性、 密码子和反密码子之间的配对很重要
第四十页,共199页。
DHU环
aa接受臂
反密码子环
TψC环
附加环
第四十一页,共199页。
(2) “ L”形三级结构
a、 “L”构型的结构力
• 二级结构中的碱基堆积力和氢键
• 二级结构中未配对碱基形成的氢键
氢键维持了 “L” 型的结
Tyr UAC, UAU GUA
Tyr UAC/UAU GUA
Lys AAA/AAG UUU
Trp UGG
CCA
CUA CUA CUA UUA UUA UCA
UAG UAG UAG UAA/UAG UAA/UAG UGA/UGG
第四十七页,共199页。
错义突变
错义突变
AUG AGA UAA
UCU
AUG GGA UAA
第四十六页,共199页。
基因
SupD(su1) SupE(su2) SupF(su3) SupC(su4) SupG(su5) SupU(su7)
表 14-6 由反密码子突变而产生的无义抑制基因
tRNA
野生型
抑制基因
识别的密码子 反密码子 反密码子 识别的密码子
Ser UCG
CGA
Gln CAG
CUG
无义抑制--也会造成对正常终止密码子的通读, 产生比野生型长的蛋白质
第四十五页,共199页。
无义突变使UUG变为UAG
Tyr-tRNA阅读UAG密码子
AUG UUG UAA AAC
AUG UAG UAA
释放因子
AUG UAG UAA
UAC
AUC
AUG
第四十页,共199页。
DHU环
aa接受臂
反密码子环
TψC环
附加环
第四十一页,共199页。
(2) “ L”形三级结构
a、 “L”构型的结构力
• 二级结构中的碱基堆积力和氢键
• 二级结构中未配对碱基形成的氢键
氢键维持了 “L” 型的结
Tyr UAC, UAU GUA
Tyr UAC/UAU GUA
Lys AAA/AAG UUU
Trp UGG
CCA
CUA CUA CUA UUA UUA UCA
UAG UAG UAG UAA/UAG UAA/UAG UGA/UGG
第四十七页,共199页。
错义突变
错义突变
AUG AGA UAA
UCU
AUG GGA UAA
第四十六页,共199页。
基因
SupD(su1) SupE(su2) SupF(su3) SupC(su4) SupG(su5) SupU(su7)
表 14-6 由反密码子突变而产生的无义抑制基因
tRNA
野生型
抑制基因
识别的密码子 反密码子 反密码子 识别的密码子
Ser UCG
CGA
Gln CAG
CUG
无义抑制--也会造成对正常终止密码子的通读, 产生比野生型长的蛋白质
第四十五页,共199页。
无义突变使UUG变为UAG
Tyr-tRNA阅读UAG密码子
AUG UUG UAA AAC
AUG UAG UAA
释放因子
AUG UAG UAA
UAC
AUC
AUG
第一、二节生物信息的传递(下)从mRNA到蛋白质1

B 、 终止密码子也与通用密码子不一样;
Codons Comparing between in usual and in m
Met
Arg
stop
Changes occur in the mitochondrial genetic code
果蝇
(3)、线粒体中密码子的特征:
U* A > G I U > C> A
二、tRNA的功能
是遗传信息从mRNA流动到蛋白质的过程中起“解码 器”的作用;
• 具有反密码子,能够识别mRNA上的密码子;
• 具有特定的三维结构,在氨酰-tRNA合成酶的帮助 下,能够和特异性氨基酸相结合;
• 由于其特定的空间构象,在核糖体中,其反密码子 臂能够和核糖体小亚基中的mRNA 相结合,另一端的 氨基酸接受臂可以使氨基酸正确插入到正在延伸的肽 链的C末端上。
I
A或U或C
摆动假说的碱基配对规律
(3)、线粒体中密码子的特征:
线粒体中具有与通用密码不同的编码信息
A、 线粒体codon较为整齐(均为2/4/6)
2 codon; F, I, Y, H, Q, N, E, k, D,W, M, C 4 codon; V, P, T, A, R, G 6 codon; L, S
(4)RNA的三级结构:L型折叠
tRNA的三级结构
• tRNA三级结构的意义:
tRNA(transfer ribonucleic asid)在蛋白质合成中处于关键地 位,它不但为每个三联体密码子译成氨基酸提供接合体,还为 准确无误地将所需氨基酸运送到核糖体上提供运送载体。
(1)被特定的氨酰- tRNA合成酶识别的结 构基础,使tRNA接受正确的活化氨基酸。 ( 2 )在蛋白质合成过程中,tRNA恰到 好处的三级结构,起着连结生长的多肽链 与核 糖体的作用。
Codons Comparing between in usual and in m
Met
Arg
stop
Changes occur in the mitochondrial genetic code
果蝇
(3)、线粒体中密码子的特征:
U* A > G I U > C> A
二、tRNA的功能
是遗传信息从mRNA流动到蛋白质的过程中起“解码 器”的作用;
• 具有反密码子,能够识别mRNA上的密码子;
• 具有特定的三维结构,在氨酰-tRNA合成酶的帮助 下,能够和特异性氨基酸相结合;
• 由于其特定的空间构象,在核糖体中,其反密码子 臂能够和核糖体小亚基中的mRNA 相结合,另一端的 氨基酸接受臂可以使氨基酸正确插入到正在延伸的肽 链的C末端上。
I
A或U或C
摆动假说的碱基配对规律
(3)、线粒体中密码子的特征:
线粒体中具有与通用密码不同的编码信息
A、 线粒体codon较为整齐(均为2/4/6)
2 codon; F, I, Y, H, Q, N, E, k, D,W, M, C 4 codon; V, P, T, A, R, G 6 codon; L, S
(4)RNA的三级结构:L型折叠
tRNA的三级结构
• tRNA三级结构的意义:
tRNA(transfer ribonucleic asid)在蛋白质合成中处于关键地 位,它不但为每个三联体密码子译成氨基酸提供接合体,还为 准确无误地将所需氨基酸运送到核糖体上提供运送载体。
(1)被特定的氨酰- tRNA合成酶识别的结 构基础,使tRNA接受正确的活化氨基酸。 ( 2 )在蛋白质合成过程中,tRNA恰到 好处的三级结构,起着连结生长的多肽链 与核 糖体的作用。
医学细胞生物学 生物信息的传递PPT文档

野生型抑制基因
识别的密码子反密码子
USer UCG CGA CA UAG Gln CAG CUG CUAUAG Tyr UAC, UAU GUACUA UAG UUA UAA/UAGTyr UAC/UAU GUA Lys AAA/AAG UUU UUAUAA/UAG Trp UGG CCAUCA UGA/UGG
20种氨基酸
核糖体较大,为80S; 新生蛋白质经蛋白酶切后变成有功能的成熟蛋白质
校正tRNA必须与正常的tRNA竞争结合密码子 在某些条件下,变性的蛋白质可能会恢复活性。 循环,每个循环包括:AA-tRNA肽与核糖体结合、 50S大亚基结合,GTP水解,释放翻译起始因子。
2×106D5S=120b 对应于同一氨基酸的密码子称为同义密码子 编码某一氨基酸的密码子越多,该氨基酸在蛋白质
普遍性与特殊性
蛋白质生物合成的整套密码,从原核生物到人类都通用。
已发现少数例外,如动物细胞的线粒体、植物细胞的叶绿体。
线粒体与核DNA密码子使用情况的比较
生物密码子
所有 酵母 果蝇
线粒体DNA编码的 氨基酸
UGA色氨酸终止子 CUA苏氨酸亮氨酸 AGA丝氨酸精氨酸
核DNA编码的氨基 酸
哺乳类 哺乳类
Tu GTP
(五)蛋白质前体的加工
N端fMet或Met的切除
新生蛋白质经蛋白酶切后变成有功能的成熟蛋白质
(四)肽链的终止
原核肽链合成终止过程
终止因子 (原核生物)
RF1:识别终止密码子
UAA和UAG
RF2:识别终止密码子
UAA和UGA
RF3:具GTP酶活性,刺激RF1和
RF2活性,协助肽链的释放
氨酰-tRNA合成酶