高分辨基因分型的临床详解演示文稿
用探针法进行基因分型的方法

用非标记探针、遮蔽技术及高分辨率熔解扩增子分析对RET原癌基因进行基因分型RET原癌基因单个碱基的突变可以引发多发性内分泌腺瘤2型。
RET突变传统的基因分型方法是外显子测序。
一种闭管操作的基因分型方法已经成熟,此方法用的是一种饱和DNA 染料,非标记探针及高分辨率熔解扩增子分析。
此方法需要两个连续的聚合酶链式反应阶段,主要的和第二次实验。
主要的实验共用7个反应和8个非标记探针分析RET基因外显子10、11、13、14和16 。
主要实验基因分型了野生型外显子,外显子13普通的多态性,外显子16的一个突变及其它检测到的序列变化。
主要非标记探针数据限制检测到的RET基因序列突变的基因分型,这些突变位点的基因分型在第二次实验的2-5个反应中进行。
设计6条探针,所用方法是:用遮蔽技术和遮蔽选择的序列变化使探针下其它位置的突变的分析变得明确。
这两步RET基因分型之后,少于实验0.2%的外显子需要测序确定基因型。
对5个野生型和29个可用的RET序列突变样品(与测序结果100%一致)做盲研究。
用非标记探针和遮蔽技术进行高分辨率熔解分析是快速、精确的基因分型方法,>50的RET序列突变可以进行基因分型。
多发性内分泌腺瘤2型(MEN2)综合症,MEN2A,MEN2B及家族性甲状腺髓样癌由生殖细胞系RET基因外显子10、11、13和16的突变引发。
MEN2综合症是常染色体显性秩序失调引起的高生命危险的甲状腺癌。
RET突变的遗传学检测可以确认处于甲状腺癌危险中但是没有爆发癌症的病人,此时切除甲状腺可以增加存活的机率。
之前有许多实验方法检测或基因分型RET突变,比如说单链构象多态性,变性梯度凝胶电泳,温度梯度毛细管电泳,限制性内切酶消化PCR产物,焦磷酸测序,荧光标记杂交探针及微卫星。
但是,这些方法中的大多数需要PCR反应之后的操作来检测突变。
其中一些方法报道突变检测的敏感性为95%或少于95%,或者仅实验了RET突变样品的一小部分。
高分辨率熔解曲线分析法之Snakback基因分型法

高分辨率熔解曲线分析法之Snakback基因分型法背景:DNA的发夹结构作为自身探针扩增用于PCR产物的分子分析。
在这之前实体分离或共价修饰的寡核苷酸荧光是必须的。
方法:我们用饱和染料LC Green做不对称PCR,40-45个循环,其中1条引物的5’端包括了一条尾巴,这条尾巴和其延伸引物互补,但这尾巴没有任何特别的共价修饰。
样品可以在LightsCycler的转子中快速的扩增,可以在96/384 模块加热PCR仪中进行高通量的扩增。
除了扩增扩增子的双链,通过引物尾巴的“弹回”及其延伸引物的杂交形成单链发夹结构。
在HR-1(毛细管)或Lightscanner(96/384孔板)上运行高分辨率熔解曲线。
结果:用弹回引物扩增的PCR产物在低温处显示两个发夹结构熔解峰,在高温处显示全部扩增子的熔解峰。
发夹结构的熔解温度与其长度(6-28bp)是线性相关,与环的大小成负相关。
我们可以轻松的将杂合突变和纯合突变进行基因分型。
用弹回探针对100个未知基因型的临床样本的F5 1691G>A进行基因分型,与之前的分型结果一致。
我们用2条弹回引物做不对称PCR,分析CFTR基因外显子10的2个范围,通过稀释产物形成分子内部杂合子进行基因分型,分出7种不同的基因型。
结论:弹回引物结合饱和染料基因分型的方法在闭管体系中,只用2条引物便可以提供一条特异的探针,此探针没有特殊的共价修饰。
在分子诊断学中,DNA的发夹结构是非常有用的工具,包括stem-loop探针和self-probing扩增。
stem-loop探针通常叫做“分子信标”,是一段共价修饰的核苷酸,一端末尾是荧光基团,另一端末尾是淬火剂。
self-probing扩增用于弹回单链构想多态性(SSCP)和蝎子引物。
弹回单链构想多态性SSCP逐渐用于在PCR产物中人工引入二级结构进行SSCP分析。
依赖于扩增序列,与引物尾巴互补的8-11bp序列弹回至其延伸产物上,形成了单链发夹结构,虽然弹回SSCP的引物没有共价修饰,电泳已经足够分离发夹结构。
高分辨染色体的识别

8号染色体 着丝粒分 化为p11.1和q11.1带。 平头整齐而浓染的p23.2 深带是识别8号染色体短 臂的一大特征。这也是 它与7号和9号染色体短 臂相区别的极重要的特 征。短臂上有12.3、 21.2、22、23.2,4条明 显的深带,长臂上各深 带分布较均匀。
9号染色体 着丝粒 紧邻p12带。短臂中 部可见21.1、23两 条明显而浓染的深带。 长臂可见分别由 21.11、21.13、 21.31、21.33,以 及31.1、31.3、33.1、 33.3所组成的两个深 带群。这也是它与8 号和10号染色体相 鉴别的主要特征。
1号染色体 p12带浓染, q12带分化为12.1、12.3两 条浓染的深带,着丝粒浓染。 短臂远端分化出36.21、 36.23两条浓染的伸带。这 是区别长短臂的主要特征, 长臂近侧段可见由12.1、 12.3、21.21、21.23、 22.1、22.3、24.1、24.3所 组成排列整齐的8条深带, 其远侧段可见由31.1、31.3、 41、43所组成的3个明显的 深带阶段。
3号染色体 着丝粒分化为p11.1和 q11.1。在短臂上可见由14.1、 14.31、14.33,3条带和22.1、 22.3、24.1、24.3,4条带所组成 的两个深带群,其间由淡染的21.1、 21.2和21.31、21.32、21.33所组 成的节段相隔,它是区别长短臂的 一个重要特征。长臂上有13.11、 13.13、13.31、13.33、21.2、 22.1、22.3、24、25.2、26.1、 26.31、26.33、27.2、28带共14 条较均匀分布的深带,其末端平头 整齐并封底的29.2淡染深带也是区 别3号染色体及其长短臂的一个可 借鉴的特征。
11号染色体 着丝粒 分化为p11.1和q11 带。短臂上可见 11.12、12、14、 15.2、15.4、这5条 分布均匀的深带,这 是它与12号染色体相 鉴别的重要特征。长 臂中段有由13.4、 14.1、14.3、22.1、 22.3这5条带组成的 深带群,末端有由 24.1、24.3所组成的 深带节段。
最新ApoE基因分型检测课件PPT

人体的ApoE基因位于第19号染色体长臂13区2带(19q13.2) 上,含有4个外显子和3个内含子。
INTERNAL USE ONLY
2. 载脂蛋白E(ApoE)简介
ApoE 基因的多态性:
ApoE由三个等位基因(ε2、ε3、ε4)相互组合形成6种主要基因型
INTERNAL USE ONLY
长 的 时 间 隧 道,袅
ApoE基因分型检测
目录
1 引言 2 载脂蛋白E(ApoE)简介 3 ApoE对心血管类疾病的风险预测 4 ApoE对阿尔茨海默病的风险预测 5 ApoE基因分型检测应用总结
INTERNAL USE ONLY
Copyright 2016 ChoutuTech LIMITED
进 入 夏 天 ,少 不了一 个热字 当头, 电扇空 调陆续 登场, 每逢此 时,总 会想起 那 一 把 蒲 扇 。蒲扇 ,是记 忆中的 农村, 夏季经 常用的 一件物 品。 记 忆 中 的故 乡 , 每 逢 进 入夏天 ,集市 上最常 见的便 是蒲扇 、凉席 ,不论 男女老 少,个 个手持 一 把 , 忽 闪 忽闪个 不停, 嘴里叨 叨着“ 怎么这 么热” ,于是 三五成 群,聚 在大树 下 , 或 站 着 ,或随 即坐在 石头上 ,手持 那把扇 子,边 唠嗑边 乘凉。 孩子们 却在周 围 跑 跑 跳 跳 ,热得 满头大 汗,不 时听到 “强子 ,别跑 了,快 来我给 你扇扇 ”。孩 子 们 才 不 听 这一套 ,跑个 没完, 直到累 气喘吁 吁,这 才一跑 一踮地 围过了 ,这时 母 亲总是 ,好似 生气的 样子, 边扇边 训,“ 你看热 的,跑 什么? ”此时 这把蒲 扇, 是 那 么 凉 快 ,那么 的温馨 幸福, 有母亲 的味道 ! 蒲 扇 是 中 国传 统工艺 品,在 我 国 已 有 三 千年多 年的历 史。取 材于棕 榈树, 制作简 单,方 便携带 ,且蒲 扇的表 面 光 滑 , 因 而,古 人常会 在上面 作画。 古有棕 扇、葵 扇、蒲 扇、蕉 扇诸名 ,实即 今 日 的 蒲 扇 ,江浙 称之为 芭蕉扇 。六七 十年代 ,人们 最常用 的就是 这种, 似圆非 圆 , 轻 巧 又 便宜的 蒲扇。 蒲 扇 流 传 至今, 我的记 忆中, 它跨越 了半个 世纪, 也 走 过 了 我 们的半 个人生 的轨迹 ,携带 着特有 的念想 ,一年 年,一 天天, 流向长
HLA-B27高分辨基因分型的临床

HLA-B27与JRA致病机制的研究 与 致病机制的研究
以各种感染性微生物作为外来抗原(HLA-B27分子与其结构 有相似性),激活免疫细胞,通过损伤或分泌细胞因子触 发异常的免疫反应,引起自身组织的损害和变性。 特别是细菌、病毒的特殊成分作为超抗原,其结构与人类 MHC抗原有同源性,不需抗原呈递细胞加工处理,即可直接 与T细胞受体直接结合(HLA-B27充当细胞表面受体),激 活T细胞。
HLA I类分子是在内质网成熟的,新合成的HLA Iα进入内质网腔 与钙连蛋白结合,使I类分子在内质网中维持部分折叠,继而与新 合成的β2-微球蛋白组装成完整的I类分子,此时B27分子才能被认 为是成熟的,转运至细胞表面。研究发现,HLA-B27折叠速度较其他 HLA分子(非HLA-B27分子)慢。导致内质网腔中出现大量的折叠不 良或部分折叠的HLA-B27重链部分,通常情况下,非折叠的重链在内 质网腔内能够被清除,但由于一些相关物质的缺乏或无效,所致非折 叠的重链堆积,形成了非折叠蛋白反应。有资料报道,AS的发生可能 与这些HLA-B27分子的未折叠有关。
Your company slogan
强直性脊柱炎(AS) 强直性脊柱炎(AS)
1973年Brewerton和schiosstein发现 强直性脊柱炎(ankylosing spondylitis AS)与HLA-B27抗原有 非常强的相关,这是目前唯一有临床 价值的血型相关抗原,这在迄今已知 的HLA与疾病的关联中是最强和最典 型的。
Your company slogan
其它疾病
【Genetic markers linked to rheumatoid arthritis are also strongly associated with articular manifestations in ulcerative colitis patients】 【Flt3一ITD mutations can generate leukaemia specific neoepitopes: potential role for immunotherapeutic approaches】 【Ankylosing spondylitis is linked to Klebsiella—the evidence】 等三篇文献报道:HLA-B27抗原与白塞氏病也 等三篇文献报道: 有一定的相关性。
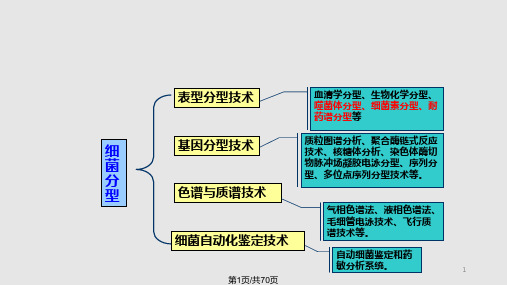
细菌的分型及其检测技术PPT课件
过夜增殖噬菌体。分离纯化单斑
3次,至平板噬斑形态大小一致
纯第化11页/共70页
11
• (三)噬菌体的增殖、保存
1、噬菌体增殖 液体增殖法:定时加入对数期菌体 固体增殖法:同时加大菌体和噬菌体的浓度 2、去除菌体
细菌滤器法和三氯甲烷法
3、噬菌体的保存
冷藏保存:无需防腐剂,分装后7d,不宜超过4w
冷冻干燥:2年
鉴定时不需活分析细菌在体外培养物中的代谢产物如厌氧菌产生的短链脂肪酸色谱图需氧菌的有机酸色谱图等临床标本中检出和鉴定细菌如检测脑脊液中的乳酸对细菌性脑膜炎做出明确诊断等热裂解气相色谱法鉴定细菌将样品干燥高温裂解碎片色谱图鉴定微生物如沙门菌属十种血清型鉴别高分辨气相色谱法分析冷冻条件下细菌的挥发性有机产物鉴定引起冷冻品腐败的微生物液相色谱法基于混合物中各组分对固定相和流动相亲和力的差异分离组分的
24
第24页/共70页
(二)培养基和抗菌药物纸片
1.抗菌药物纸片:商品化,选择直径为,吸水量为20μl的专用药敏纸片,用逐片 加样或浸泡方法使每片含药量达规定要求。含药纸片密封贮存于超低温冰箱。
2.培养基:水解酪蛋白(M-H)培养基是兼性厌氧菌和需氧菌药敏试验标准培养 基,4℃保存。
25
第25页/共70页
16
第16页/共70页
第二节 细菌素分型技术
一、概述
• 细菌素(bacteriocin)
• 是某些细菌产生的蛋白质类抗菌物质,这种物质仅 对与产生菌亲缘关系近的细菌有抗菌作用,而对产 生菌本身不敏感。如大肠菌素、绿脓菌素、蜡样芽 胞杆菌素等。
• 抗生素
• 抗菌素是细菌、真菌等微生物在生长过程中为了生 存竞争需要而产生的化学物质,这种物质可保证其自 身生存,同时还可杀灭或抑制其它微生物。细菌产生 的有多粘菌素、抗菌肽等。
高通量宏基因组测序技术检测病原微生物的临床应用规范化专家共识ppt课件

鼓励微生物学、遗传学、临床医学等相关领域的专家加强跨学科合作, 共同推动高通量宏基因组测序技术在病原微生物检测中的临床应用和研 究进展。
THANKS FOR WATCHING
感谢您的观看
样本质量
01
样本采集和处理环节对测序结果至关重要,需要严格
控制样本质量,避免污染和误差。
数据分析
02 高通量测序产生的数据量庞大,需要建立完善的数据
分析流程和标准,确保结果的准确性和可靠性。
技术更新
03
随着测序技术的不断发展和进步,需要保持对新技术
的关注和应用,不断提高检测水平和效率。
06 总结和展望
02 高通量宏基因组测序技术 在实验室的应用
样本收集和处理
样本选择
选择适当的临床样本,如血液、 尿液、呼吸道分泌物等,根据患 者的症状和疑似病原体进行针对
性收集。
样本处理
对收集到的样本进行适当的处理 ,如离心、过滤等,以去除杂质
和富集病原微生物。
核酸提取
采用合适的核酸提取方法,如磁 珠法、柱层析法等,提取样本中
未来高通量宏基因组测序技术在病原微生物检测中的潜力 和前景
技术迭代升级
随着技术的不断发展,未来高通量宏基 因组测序技术的检测精度、效率和成本 等方面将持续优化,为病原微生物检测 提供更加可靠的技术支持。
VS
多组学联合分析
宏基因组测序技术可以与其他组学技术( 如代谢组学、蛋白质组学等)进行联合分 析,深入挖掘病原微生物与宿主之间的相 互作用机制,为临床诊断和治疗提供更加 全面的信息。
05 案例分析和经验教训
案例介绍
案例背景
01
介绍某疫情的情况,包括疫情规模、影响范围、病原微生物特
tpmt基因分型

tpmt基因分型TPMT基因分型与药物代谢相关的研究已经引起了广泛的关注。
TPMT 基因编码的酶是一种重要的药物代谢酶,参与对硫唑嘌呤、6-巯基嘌呤以及一些抗癌药物的代谢。
不同基因型的个体对于药物的代谢能力有所不同,因此TPMT基因分型也被用作个体化用药的参考依据。
TPMT基因的分型主要包括三种:TPMT*1、TPMT*2和TPMT*3。
其中,TPMT*1是正常型,即正常的酶活性;TPMT*2和TPMT*3则是突变型,这两种突变型基因会导致酶活性降低或完全丧失。
根据不同的基因型,个体的药物代谢能力也存在差异,从而影响药物的疗效和安全性。
TPMT基因的分型对于临床用药有着重要的指导意义。
研究发现,TPMT*3A是最常见的突变型基因,约占白人的10%和亚洲人的3%。
携带TPMT*3A基因的患者对于硫唑嘌呤和6-巯基嘌呤的代谢能力明显降低,容易出现药物的毒副作用。
因此,在给这类患者使用这两种药物时,需要根据其基因分型来调整药物剂量,以避免不良反应的发生。
除了TPMT*3A以外,还有其他一些较为罕见的突变型基因,如TPMT*2和TPMT*3C。
这些基因的突变也会导致酶活性的下降,从而影响药物的代谢。
因此,在用药前对患者进行TPMT基因的检测,可以帮助医生评估患者的药物代谢能力,从而制定个体化的用药方案。
TPMT基因分型在临床上的应用不仅限于对硫唑嘌呤和6-巯基嘌呤的用药调整,还可以用于其他一些抗癌药物的个体化用药。
例如,6-巯基嘌呤是一种常用的抗癌药物,其在体内的代谢过程中需要依赖TPMT酶的参与。
因此,对于患有突变型TPMT基因的患者,使用6-巯基嘌呤时需要减少剂量或选择其他的治疗方案,以避免药物的毒副作用。
除了对药物代谢的影响外,TPMT基因分型还与一些自身免疫疾病的发生有关。
研究发现,携带TPMT*3C基因的个体在使用硫唑嘌呤等药物时,容易出现骨髓抑制等不良反应,进而增加自身免疫疾病的风险。
因此,在给这类患者用药时,需要谨慎选择药物并调整剂量,以减少不良反应的发生。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
HLA-B*2705基因分布最广,几乎见于所有人种。 甚至认为其它等位基因都是由HLA-B*2705基因发 生突变、移位形成的。 HLA-B*2705编码的蛋白最为显著的特点就是在B袋 上的45位发生氨基酸的突变,这一改变直接影响 HLA-B27分子在细胞表面的表达,影响HLA-B27分 子与抗原肽的结合,同时影响了HLA-B27分子对胞 内外抗原的递呈。
HLA-B27分子游离重链与AS
HLA-B27分子的主要功能是:识别和处理抗原肽,然后在 递呈给T淋巴细胞。HLA-B27分子具有特殊的生物学效应, 它在体外具有形成重链二聚体的能力,主要是通过B27分 子第67位氨基酸的残基半氨酸的二硫键结合形成稳定的二 聚体。通过认识HLA-B27分子结构的特异性,重要的是深 入研究游离重链的表达是否应该认为是导致AS的发病原因 之一。当HLA-B27分子适当的、正确的组装时,其递呈的 抗原肽通常被CD8 CTL识别;但当AS发生时,限制性的 CIM Th细胞却能识别非折叠、空载或缺失的B27分子。在 这种情况下,就是HLA II类分子递呈B27重链,降解后形 成的多肽被CD4 Th细胞识别后导致关节炎症。此研究目 前还存在争议。
【differential diagnosis juvenile ankylosing spondylitis and juvebile rheumatoid arthritis by subtypes of
不同人群AS患者的HLA-B*27等位基因频率
摘自【HLA—B*27高分辨等位基因在1 606例疑似强直性脊柱炎患者中的表达】
HLA-B27与AS致病机制的研究
HLA-B27分子与非折叠蛋白理论假说 HLA-B27分子游离重链与AS 其它
HLA-B27分子非折叠蛋白理论假说
HLA I类分子是在内质网成熟的,新合成的HLA Iα进入内质网腔 与钙连蛋白结合,使I类分子在内质网中维持部分折叠,继而与新 合成的β2-微球蛋白组装成完整的I类分子,此时B27分子才能被认 为是成熟的,转运至细胞表面。研究发现,HLA-B27折叠速度较其他 HLA分子(非HLA-B27分子)慢。导致内质网腔中出现大量的折叠不 良或部分折叠的HLA-B27重链部分,通常情况下,非折叠的重链在内 质网腔内能够被清除,但由于一些相关物质的缺乏或无效,所致非折 叠的重链堆积,形成了非折叠蛋白反应。有资料报道,AS的发生可能 与这些HLA-B27分子的未折叠有关。
其中HLA-B*2701与HLA-B*2705编码的蛋白 在78-81范围内有2个氨基酸的不同; 其中HLA-B*2702与HLA-B*2705编码的蛋白 在78-81范围内有3个氨基酸的不同; 其中HLA-B*2703与HLA-B*2705编码的蛋白 仅有1个氨基酸的差别,即在59位前者 为组氨酸(His),后者为酪氨酸(Tyr);
其它
HLA-B27可提高致关节炎的细菌在胞内的存活率, 从而加剧关节炎的严重度。 B27改变信号转导学说,即HLA-B27通过修饰 炎症因子基因相关的信号转导过程而致关节炎。 HLA-B39,HLA-B60可能对AS的诱发起到一定的作用。
幼年类风湿性关节炎(JRA)
幼年类风湿性关节炎(juvenile rheumatoid arthritis JRA)是类风湿性关 节炎疾病中的一个特殊类型,而HLA-B27抗 原阳性的幼年类风湿关节炎又是类风湿关节 炎疾病中的一个亚型。幼年类风湿性关节炎 是儿童时期较常见的结缔组织病,国外的报 道发现,较多的幼年类风湿性关节炎患者的 HLA-B27抗原呈现阳性,且没有男女性别差 异。
高分辨基因分型的临床详解演 A-B27抗原分子及其等位基因
HLA-B27抗原分子结构及其特点
HLA-B27抗原分子是由位于人类第六号染色 体短臂上B*27等位基因编码的糖蛋白, 分子量 为56×103U。 由两条多肽链组成,一条为α链或重链,分子 量约为44×103U,包括胞外区,跨膜区,胞 内区三个结构域,其中胞外区又分为α1,α2, α3三个结构区;另一条为β链或轻链即β2微 球蛋白,分子量约12×103U,二者结合共同 组成HLA-B27抗原分子。
HLA-B27抗原分子结构
α 1区与α2区的肽链折叠后共同构成一个抗原 结合凹槽,槽内形成A,B,C,D,E,F共6个袋状结 构,其中B袋在HLA-B27分子中是最保守的结构。 凹槽的作用是结合抗原肽,使抗原肽与HLA-B27 分子牢固的结合。
HLA-B*27等位基因结构及其特点
2002年WHO正式命名的HLA—B*27 等位基因达到了24个,其中只有15个 有相对应的抗原表达。到2009年11月 WHO正式命名HLA—B*27等位基因 达到59个(B*2701一B*2759) 。 HLA—B*27等位基因呈多态性,主要 是因为个别的碱基突变造成的,编码蛋 白的差异主要集中在αl、α2结构区内。
AS的抗原学检测
【HLA-B27与强直性脊柱炎的临床分析】报 道:HLA-B27抗原阳性率在AS患者高达96% 以上,而正常群体中仅4%-7%,HLA-B27抗原 携带者发生AS的相对危险率高达70%。
与AS相关的B*27等位基因
AS的基因学检测
不同人群的AS患者涉及的B*27等位基因
与AS相关的B*27等位基因
2.与HLA-B27相关的疾病
强直性脊柱炎 (ankylosing spondylitis AS)
幼年类风湿关节炎 (juvenile rheumatoid arthritis JRA)
急性前葡萄膜炎 (acute anterior uveitis,AAU) 白血病等疾病
强直性脊柱炎(AS)
1973年Brewerton和schiosstein发现 强直性脊柱炎(ankylosing spondylitis AS)与HLA-B27抗原有 非常强的相关,这是目前唯一有临床 价值的血型相关抗原,这在迄今已知 的HLA与疾病的关联中是最强和最典 型的。