第11章 酶作用机制和酶活性调节
合集下载
酶活性的调节

• 与共价修饰一样,水解激活是也一种全或无的调节方 式,酶原状态没有活性,但与共价修饰不同的是,它 是不可逆的,即一旦被激活就不可能回到原来的非活 性的酶原状态。
胰蛋白酶的水解激活
几种蛋白酶的水解激活
糜蛋白酶的水解激活
某些凝血因子的水解激活
调节蛋白的激活或抑制
• 某些蛋白质能够作为配体与特定的酶结 合而调节被结合酶的活性,这些调节酶 活性的蛋白质称为调节蛋白,其中,激 活酶活性的调节蛋白称为激活蛋白,抑 制酶活性的蛋白称为抑制蛋白。抑制蛋 白通常结合在酶的活性中心阻止底物与 活性中心结合而达到抑制的效果。
乳酸脱氢酶催化的酶促反应
酶的“质变”
• 别构调节、共价修饰、水解激活、调节蛋白的结 合和解离以及单体的聚合和解离。
酶活性的别构调节、共价修饰和水解激活调节的异同
性质 别构调节
共价修饰
水解激活
可逆性 是
是
否
全或无 否
是
是
酶调节 否 是(蛋白质激酶和 是(蛋白酶) 磷蛋白磷酸酶)
别构调节
• 别构调节的原理在于一些酶除了活性中心以外,还含有所谓的别构中心, 该中心能够结合一些特殊的配体分子(有时为底物)。当别构中心结合配 体以后,酶构象发生改变,从而影响到活性中心与底物的亲和力,并最终 导致酶活性发生变化。能够进行别构调节的酶称为别构酶,与别构中心结 合调节酶活性的配体分子称为别构效应物。起抑制作用的别构效应物称为 别构抑制剂,起激活作用的别构效应物称为别构激活剂。由底物作为别构 效应物产生的别构效应称为同促效应,否则,就称为异促效应。许多别构 酶具有多个别构中心,能够与不同的别构效应物结合。
• 具有协同效应的酶和无协同效应的酶都可以受到别构调节。
酶活性的反馈抑制
胰蛋白酶的水解激活
几种蛋白酶的水解激活
糜蛋白酶的水解激活
某些凝血因子的水解激活
调节蛋白的激活或抑制
• 某些蛋白质能够作为配体与特定的酶结 合而调节被结合酶的活性,这些调节酶 活性的蛋白质称为调节蛋白,其中,激 活酶活性的调节蛋白称为激活蛋白,抑 制酶活性的蛋白称为抑制蛋白。抑制蛋 白通常结合在酶的活性中心阻止底物与 活性中心结合而达到抑制的效果。
乳酸脱氢酶催化的酶促反应
酶的“质变”
• 别构调节、共价修饰、水解激活、调节蛋白的结 合和解离以及单体的聚合和解离。
酶活性的别构调节、共价修饰和水解激活调节的异同
性质 别构调节
共价修饰
水解激活
可逆性 是
是
否
全或无 否
是
是
酶调节 否 是(蛋白质激酶和 是(蛋白酶) 磷蛋白磷酸酶)
别构调节
• 别构调节的原理在于一些酶除了活性中心以外,还含有所谓的别构中心, 该中心能够结合一些特殊的配体分子(有时为底物)。当别构中心结合配 体以后,酶构象发生改变,从而影响到活性中心与底物的亲和力,并最终 导致酶活性发生变化。能够进行别构调节的酶称为别构酶,与别构中心结 合调节酶活性的配体分子称为别构效应物。起抑制作用的别构效应物称为 别构抑制剂,起激活作用的别构效应物称为别构激活剂。由底物作为别构 效应物产生的别构效应称为同促效应,否则,就称为异促效应。许多别构 酶具有多个别构中心,能够与不同的别构效应物结合。
• 具有协同效应的酶和无协同效应的酶都可以受到别构调节。
酶活性的反馈抑制
酶的作用机制和酶的调节101214

酶的别构调节:酶分子的非催化部位与某些化合物可逆地非
共价结合后发生构象的改变,进而改变酶的活性状态。
别构酶:具有别构调节作用的酶。 效应物(别构剂):凡能使酶发生别构作用的物质。通常为
小分子代谢物或辅因子。
正效应物(别构激活剂):因别构导致酶活性增加的物质。
负效应物(别构抑制剂):因别构导致酶活性减少的物质。
⑴ 疏水环境(酶分析表面的裂缝)
介电常数低,加强极性基团间的作用。
⑵ 电荷环境
在酶活性中心附近,往往有一电荷离子,可稳定过渡态的离子。
四、 酶催化反应机制的实例(自学)
(一)溶菌酶(lysozyme)(P394)
溶菌酶存在于蛋清和动物的眼泪中,其生物学功能是催化某些 细菌细胞壁多糖的水解,从而溶解这些细菌的细胞壁。
三、 影响酶催化效率的有关因素
(一)底物和酶的邻近效应与定向效应(P388)
1.邻近效应(approximation) :指酶与底物结合成ES后,使底
物和底物之间(如双分子反应) 、酶催化基团与底物
之间结合于同一分子而使有效浓度得以极大升高, 从而使反应速率大大增加的一种效应。
2.定向效应(orientation) :指反应物的反应基团之间、酶的催
也可能在不同亚基上。
每个别构酶分子可以有一个以上的活性部位和调节部位,因此 可以结合一个以上的底物分子和调节物分子。
T(tense) - 态
R(relax) -态
PALA(N-磷乙酰-L-天冬氨酸)结合到ATCase活性中心的模型
图10-52
(5)底物结合到ATCase上引起高度协同的别构转变。
(6)ATP和CTP通过改变T态和R态之间的平衡来调节 ATPcase 的活性。
第十一章酶的催化机理和活性调节

与酶的高效性有关的因素
a.底物与酶的靠近与定向
b.酶使底物的敏感键发生变形
c.共价催化 d.酸碱催化 e.低介电区域的活性中心 f.金属离子催化作用
108
103
g.多功能催化作用
不同的酶,引起其高效性的因素是不同的, 可以受一种或几种因素的影响
第11章
酶的作用机制和酶的调节
第十一章
酶的作用机制和酶的调节
一、酶的活性中心 二、酶的催化机理 三、丝氨酸蛋白酶类 四、酶活性的调节 五、同工酶
(一) 酶分子的结构特点
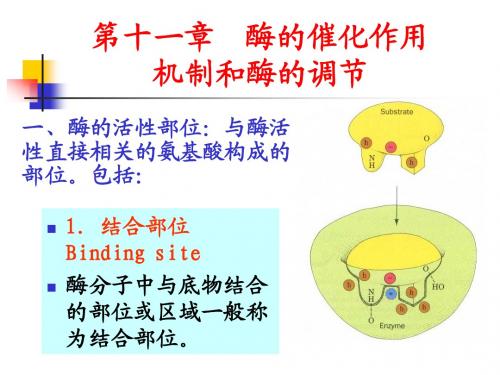
• 1.结合部位 Binding site • 酶分子中与底物结合 的部位或区域一般称 为结合部位。
2.催化部位 catalytic site
• 酶分子中促使底物发生化 学变化的部位称为催化部 位。 • 结合部位决定酶的专一性, • 催化部位决定酶所催化反 应的性质与能力。
径的第一步反应的酶相结合,结合的结果使这个 酶活性下降,从而使整条代谢途径的反应速度慢 起来。这种情况又称为“反馈抑制 ”。
正反馈
结合使酶活性增强
第一个酶
(有活性)
终产物
第一个酶
(无活性)
终产物(调节物) 结合在调节中心
(一)变构调节(别构调节):
某些代谢物能与变构酶分子上的变构部位 特异性结合,使酶的分子构象发生改变, 从而改变酶的催化活性以及代谢反应的速 度,这种调节作用就称为变构调节 (allosteric regulation)。 调控部位:酶分子中存在着一些可以 与其他分子发生某种程度的结合的部 位,从而引起酶分子空间构象的变化, 对酶起激活或抑制作用。
2+
O P O O
O P O O CH2 H H OH O A H H OH
11 酶的作用机制和调节-王镜岩生物化学(全)

三、酸碱催化
• 酸碱催化是通过瞬时的向反应物提供质子或从反 应物接受质子以稳定过渡态,加速反应的一类催 化机制。
• 组氨酸的咪唑基的重要意义(许多酶的活性中心都有组 氨酸残基)
四、共价催化
• 共价催化:在催 化时,亲核基团 或亲电基团发生 亲核取代和亲电 加成反应, 通过共
价键与底物形成不 稳定的共价酶 - 底物 复合物 , 降低反应 :
糖原磷酸化酶
各种类型可逆的共价修饰
ATP结构式
ATP的形成
尿苷酸结构式
第四节 同 工 酶
1. 同工酶定义
• 同工酶是指催化相同的化学反应,但其 蛋白质分子结构、理化性质和免疫性能 等方面都存在明显差异的一组酶。 • 存在部位:同工酶不仅存在于同一个体 的不同组织中,甚至同一组织、同一细 胞的不同亚细胞结构中。
6、底物通过次级键结合到酶上。
酶与底物结成ES复合物主要靠次级键: 氢键、盐键、范德华力和疏水相互作用
7、 酶活性部位相对于整个酶分子来
说更敏感 (变性时首先失活)
二、研究酶活性部位的方法
• 1.侧链基团的化学修饰法 • 可以被化学修饰的基团很多,如巯基、 羟基、咪唑基、氨基、羧基和胍基等。
• 2.定点诱变法
大增加了底物的有效浓度)
•定向效应是指反应物的反应 基团之间或酶的催化基团与底 物的反应基团之间的正确取位。
二、底物的形变和诱导契合
• 当酶遇到其专一性底 物时,酶中某些基团 或离子可以使底物分 子内敏感键的某些基 团的电子云密度增高 或降低,使敏感键的 一端更加敏感,底物 分子发生形变,底物 比较接近它的过渡态, 降低了反应活化能, 使反应易于发生。
酶催化机理的实例
胰凝乳蛋白酶(电荷中继网) 催化三联体 Asp---His---Ser
第十一章 酶的作用机制和酶活性的调节

第十一章酶的作用机制和酶活性的调节
班级:姓名:学号:成绩:
一、选择题
1.有机磷农药,作为酶的抑制剂,作用于酶活性中心的答()
①巯基;②羧基;
③羟基;④氨基。
2.保护巯基酶免遭氧化而失活的化合物是答()
①胱氨酸;②二价阳离子;
③谷胱甘肽;④尿素
3.羧肽酶催化水解释放肽链中含下列哪种自由基团的氨基酸?答()
①ε氨基;②α羧基;
③β羧基;④γ羧基
4.胰蛋白酶催化断裂的肽键要求羧基参与成键的氨基酸是答()
①碱性氨基酸;②酸性氨基酸;
③中性氨基酸;④任一种氨基酸
二、名词解释
1.邻近效应:
2.激酶(kinases):
3.激活作用:
4.广义(一般)酸碱催化:
5.亲核催化:
6.共价催化:
7.巯基酶:
8.活性部位(活性中心)
9.亲和标记
10.催化三联体(电荷中继网)
11.别构酶
12.共价修饰
13.酶原激活
14.同工酶
三、问答题(共15分)
1.简述对酶的催化能力作出贡献的因素。
2.确定酶的活性部位主要采用哪些方法?
3.设计一个实验,以确定酶的一个残基是否为该酶活性所必需的。
4.关于同工酶的下列说法,哪一些是错误的?并简要分析
(1)在诊断心肌梗死中是重要的。
(2)在诊断肝病方面是重要的。
(3)有组织专一性
(4)它在临床上的价值决定于个体之间的遗传差异。
酶的作用机制和酶的调节
O H2N CH C OH CH CH2 O C O OH O CH CH2 CH2 CH2 CH2 CH2 NH2 OH OH C H2N OH CH C OH NH2 O O C OH COOH
CH2 H2N CH2 C OH
酸、碱性基团: 天冬氨酸和谷氨 酸的羧基,酪氨 酸的酚羟基 赖氨酸的氨基, 组氨酸的咪唑基。
酶分子侧链基团的化学修饰法 非特异性共价修饰 特异性共价修饰 亲和标记 动力学常数测定法 X射线晶体结构分析法 定点诱变法
二、酶催化反应的独特性质
三、影响酶催化效率的有关因素
酶是专一性强, 催化效率很高的 生物催化剂,这 是由酶分子的特 殊结构决定的。 多种因素可以使 酶反应加速:
1、底物和酶的邻近效应和定向效应
5、 金属离子催化作用
*根据金属离子与酶蛋白结合程度,可分为两类: 金属酶和金属—激活酶。 (1)金属酶:酶蛋白与金属离子结合紧密。如 Fe2+/ Fe3+ 、Cu+/Cu3、Zn2+ 、Mn2+、Co2 等过 渡态离子。 (2)金属—激活酶:金属离子与酶的结合一般 较松散。在溶液中,酶与这类离子结合而被激 活。如Na+ 、K+、 Mg2+、 Ca2+ 等。 以3种主要途径参加催化过程: (1)通过结合底物为反应定向
(2)通过可逆改变金属离子 氧化态调节氧化还原反应
许多氧化-还原酶 中都含有铜或铁离 子,它们作为酶的 辅助因子起着传递 电子的功能。 Fe3+ e Fe2+ e Cu2+ Cu+
(3)通过静电稳定或屏蔽负电荷
电荷屏蔽作用是酶中金属离子的一个重要功能。
酶作用机制及酶调节

胰核糖核酸酶A
胰核糖核酸酶A
胰核糖核酸酶 A
胰 核 糖 核 酸 酶
A
羧肽酶A
丝氨酸蛋白酶族
消化作用的丝氨酸蛋白酶
• 胰蛋白酶、胰凝乳蛋白酶和弹性蛋白酶是一组 密切相关的水解酶类,它们的作用是水解肽链。
• 在胰脏内合成的是它们没有活性的酶原,然后 被分泌到消化道,并且仅仅在使用前被活化。 这3种酶各有分工,每种酶在不同类型氨基酸 侧链相邻的肽键处水解蛋白质链。
位点被10%饱和时的底物浓度 Rs≈81 米氏酶 Rs>>81 别构酶,负协同效应 Rs<<81 别构酶,正协同效应
通过n值(协同系数)
n≈1 米氏酶 n>>1 别构酶,正协同效应 n<<1 别构酶,负协同效应
别构效应的生理意义:酶对底物量的变化十分敏 感。
比如:对米氏酶而言,[S]90%Vm/[S]10% Vm=81,意思是[S]提高了81倍,v才提高9 倍,说明酶对[S]的变化很迟钝。 而对于一般的别构酶而言,[S]90%Vm/[S] 10%Vm=3,意思是[S]只要提高了3倍,v 就能提高9倍,说明酶对[S]的变化很敏感。
变构抑制剂
别构酶(Allosteric enzyme)的特点:
➢ 1)寡聚酶,由多亚基组成,包括催化部位和调节 (别构)部位;
➢ 2)具有别构效应。别构中心结合了效应物(效应 物)后,导致酶的构象发生改变,影响了活性中 心对底物的催化作用
根据别构物性质
相同(均为底物):同促效应(homotropic effect) 不同(效应调节物):异促效应
(1)临近定向效应
在酶促反应中,底物分子结合到酶的活性中心,一方 面底物在酶活性中心的有效浓度大大增加,另一方面, 由于活性中心的立体结构和相关基团的诱导和定向作 用,使底物分子中参与反应的基团相互接近,并被严 格定向定位,使酶促反应具有高效率和专一性特点。
酶的作用机制和酶活性的调节.ppt

O
O
HH
HH33CC
CC OO
OO CC
OO--
+O
O H HH NH
OC HH C HH H
几种常见酶的结构与功能
蛋白酶
• 蛋白酶是催化肽键水解的一类酶的总称。尽管肽键的水解在能量学上 是十分有利的,但如果没有蛋白酶的催化,一个肽键在中性pH和25℃ 条件下大概需要300年~600年的时间才能完成水解。
四面体中间物因肽氧负离子与Ser195以及Gly193之间形成的氢键而得 到稳定,这些相互作用的净效应导致活化能的降低。
肽键断裂,离开基团(原来肽键的氨基一侧作为第一产物)从His57 咪唑环上得到一个质子。原来肽键的羧基一侧通过氢键以及与Ser195形成 的共价键仍然与酶结合。
水进入活性中心,质子化His57使之成为酸。而释放出来的OH-亲核 进攻留下来的多肽羰基碳,于是第二个四面体形的过渡态中间物形成了 。
广义的酸碱催化
溶菌酶Glu35的广义酸催化溶菌酶Biblioteka Glu 35被疏水氨基酸残基所环绕
核糖核酸酶A的广义酸碱催化
静电催化
• 活性中心电荷的分布可用来稳定酶促反应的过 渡态,酶使用自身带电基团去中和一个反应过 渡态形成时产生的相反电荷而进行的催化称为 静电催化。
• 有时,酶通过与底物的静电作用将底物引入到 活性中心。
乳酸脱氢酶催化的酶促反应
What is an isozyme?
(1) Isozymes are physically distinct forms of the same enzyme.
(2) Isozymes may differ from each other by differences in their amino acid sequences or by the presence of different posttranslational modifications in each isozyme.
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
• 定向效应:当专一性底物向酶活性中心靠近时,会 诱导酶分子构象发生改变,使酶活性中心的相关基 团和底物的反应基团正确定向排列。
邻近定向效应
2.诱导契合和底物的形变
当酶与底物靠近时,不仅酶构象受底物 作用而变化,底物分子也常常受酶作用而 变化,也就是酶使底物分子中的敏感键发 生“变形”,从而促使底物中的敏感键更 易于破裂。
但是Asp-His-Ser组成的活性位点的排列是相似的。
32
胰蛋白酶、胰凝乳蛋白酶、弹性蛋白 酶的比较
胰蛋白酶:Arg-X, Lys-X 胰凝乳蛋白酶: 芳香族氨基酸 Phe-X, Tyr-X, Trp-X 弹性蛋白酶:Ala-X, Gly-X, Ser-X, Val-X(丙甘撕缬)
原因:酶与底物结合部位,即“口袋”,在结构上的 微小差异引起的。
2、催化基团 催化基团决定酶所催化反应的性质,同时 也是决定反应的高效性。
3、调控基团 对酶起激活或抑制作用的基团。
酶的活性中心示意图
胰蛋白酶活性位点
7
酶活性部位的特点(P172)
1. 只占酶分子总体相当小的部分; 2. 是一个三维实体; 3. 活性部位与底物分子经诱导契合互补; 4. 位于酶分子表面的一个裂缝内; 5. 底物通过较弱的次级键与酶结合; 6. 活性部位具有柔性或可运动性。 7. 酶活性中心的基团主要有:
催化三联体。
His的咪唑基是Asp羧 基和Ser羟基间的桥梁, 形成电荷中继网。
30
胰蛋白酶、胰凝乳蛋白酶、弹性蛋白 酶的比较
结构(一级结构和三维结构)及机理:极其相似
31
催化三联体具有相似的结构
枯草杆菌蛋白酶
胰凝乳蛋白酶
枯草杆菌蛋白酶和胰凝乳蛋白酶结构有很大的不同,而且组成
催化三联体的残基处于一级顺序中不同的位置。
速率也不会发生大的改变。 保证糖酵解的正常进行。
54
五、酶活性共价修饰调节
(一)酶的可逆共价修饰
蛋白质的磷酸化与脱磷酸化过程,是生物体内一 种普遍的调节方式。
由蛋白激酶和蛋白磷酸酶分别催化完成,以ATP 作为磷酸基供体。
底物蛋白质被磷酸化的氨基酸残基主要是Ser、 Thr,个别为Tyr。
可转变成有活性的酶。 酶原中,酶的活性部位被掩盖或者没有形成,酶原
的激活实质上就是酶活性部位形成的过程。
57
内肽酶 胃蛋白酶 胰蛋白酶 胰凝乳蛋白酶 弹性蛋白酶 外肽酶 羧肽酶 氨肽酶
1. 消化系统中酶原激活
消化系统中的酶多以酶原的形式合成和贮存。 酶原在一定条件被激活。
55
磷酸化酶 a(有活性)磷 酸 化酶 磷 酸 酶 磷酸化酶 b(无活性) 磷酸化酶 激 酶
葡萄糖 n Pi 葡萄糖 n1 1 磷酸葡萄糖
56
五、酶活性共价修饰调节 (二)酶原的激活-不可逆共价调节
酶原(无活性的前体酶)(有活性)
酶原:没有活性的酶的前体。 酶原的激活:酶原在一定条件下经适当的物质作用
48
(四)别构酶举例 1. 天冬氨酸转氨甲酰酶(ATCase)
1) 氨甲酰磷酸 2) 两种底物与酶的结合具有正协同效
应,表现为S型的动力学曲线。 正调接物:ATP,作为信号表明有
能量供DNA复制使用并引发需求嘧 啶核苷酸的生物合成 负调节物:CTP,在嘧啶核苷酸已 丰足时,不再进行不必要的氨甲酰 天冬氨酸及其他后续中间产物的合 成
正协同性(同促)相互作用:酶的一个亚基上活性部 位与底物(调节物)结合,增加其余亚基活性部位的亲 和力。 负协同性(异促)相互作用:酶的一个亚基上活性部 位与底物(调节物)结合,降低其余亚基活性部位的亲 和力。
44
(二)别构酶的动力学特征
12 3
P184 图11-16A
1. 无调节物:双曲线 2. 正协同作用: S形曲线 3. 负协同作用:表观双曲线
或是通过质子碱接受部分质子,达到降低反 应活化能的目的的一种作用。 酶参与的一般都是广义的酸-碱催化方式。
19
酶分子中可以作为广义酸、碱的基团
广义酸碱催化:质子供体及质子受体的催化
20
2. 共价催化 P174
催化剂与底物之间瞬间形成共价键以提高反 应速率的一种催化机制。
包括亲核催化和亲电催化。 催化剂通过放出电子或获取电子并作用于底
定酶的专一性;
催化部位 catalytic site:负责催化底物键的断裂 形成新键,决定酶的催化能力和催化反应性质。
3
酶的活性中心是指结合底物和将底物转 化为产物的区域,通常是相隔很远的氨 基酸残基形成的三维实体。
活性中心
结合基团 催化基团 调控基团
中心内必需基团 中心外必需基团
1、结合基团 酶的结合基团决定酶反应的专一性。
P188 图11-21
50
天冬氨酸转氨甲酰酶构象的转变
2个催化3聚体 3个调节2聚体
ATP
CTP P189 图11-22
51
(四)别构酶举例 天冬氨酸转氨甲酰酶(ATCase)
天冬氨酸蛋白酶的结构特点:每一种酶的活 性部位具有两个天冬氨酸残基。
天冬氨酸蛋白酶的作用机制
52
(四)别构酶举例 2. 甘油醛-3-磷酸脱氢酶
的底物(对甲苯磺酰苯丙氨酸乙酯)类似物。
P195: 3
胰蛋白酶的亲和标记试剂
O CH2 CH C CH2Cl
NH SO2 (CH2)4 NH3+ N-(ε -氨基-丁基)磺酰--L-苯丙氨酰氯甲基酮
苯丙氨酸乙酯
丝氨酸蛋白酶底物结合部位:
胰凝乳蛋白酶-芳香环
胰蛋白酶-带正电荷的Lys、Arg进入
胰蛋白酶的口袋底部有一个负电荷的Asp189,能促 进与带正电荷的Arg, Lys结合。
弹性蛋白酶的口袋较浅,只能容纳侧链较小的残基。
33
酶与底物结合部位,即“口袋”,在 结构上有微小差异
P180 图11-12
34
胰凝乳蛋白酶反应的详细机制(1)
底物
结合底物
11
His57 质子供体
23
3-磷酸甘油醛
1,3-二磷酸甘油酸
53
(四)别构酶举例 2. 甘油醛-3-磷酸脱氢酶
负协同效用。有4个亚基,可以和4个[NAD+]结 合。 [NAD+]的结合引起该亚基的构象发生变化, 并降低了同一酶分子中相邻的空位亚基的结合 亲和力。K1<K2<K3<K4。
生理意义: 在[NAD+]浓度很低时,能保证与酶的结合; 在[NAD+]浓度很高时,由于负协同效应,反应
酶
底物 同时形变
产物
底物分子发生形变
三、酶促反应机制的举例
(一)丝氨酸蛋白酶 1. 结构特点: 丝氨酸蛋白酶家族包括: 胰蛋白酶、胰凝乳蛋白酶、弹性蛋白酶、 凝血酶等,前三种酶为消化酶。 丝氨酸残基是酶活性必需的。
29
2. 丝氨酸蛋白酶的作用机制
丝氨酸蛋白酶的三个 极性残基- Ser-195, His-57, and Asp-102
P185 图11-17
P185 图11-18
47
2. KNF模型/序变模型
当配体不存在时,别构酶以T型构象存在,结合配体后由T型向R型 转变;
别构酶以序变方式进行构象变化。 亚基间的相互作用可能是正协同效应,也可能是负协同效应。 此序变模式可以较好的说明非底物调节的效应。
P187 图11-20
23
(二)酶具有高催化能力的原因
1. 底物和酶的邻近效应与定向效应 2. 诱导契合和底物形变 3. 电荷极化和多元催化 4. 疏水的微环境的影响
24
1、 邻近效应、 定向效应
• 邻近效应:在酶促反应中,由于酶和底物分子之间 的亲和性,底物分子有向酶的活性中心靠近的趋势, 使底物在酶活性中心的有效浓度大大增加的效应。
第 十一 章 酶作用机制和酶活性调节
1
主要内容
1. 酶的活性部位及其确定方法(重点难点考点) 2. 酶促反应机制(掌握) 3. 酶促反应机制的举例(重点、考点) 4. 酶活性的别构调节(重点、考点) 5. 酶活性的共价调节(重点) 6. 同工酶(了解)
2
一、酶的活性部位及其确定方法
酶的活性部位:与酶活力直接相关的区域 结合部位 binding site:负责与底物的结合,决
45
(三)协同性配体结合的模型
亚基之间存在协同效应 当一个亚基结合底物或效应物后,该亚基的构
象发生变化,从而影响其他亚基也发生构象变 化。 别构模型:
齐变模型/MWC模型/对称模型 序变模型
46
1. MWC模型/齐变模型/对称模型
每个亚基对一种配体只有一个结合位点; 每种亚基有两种构象, R型 T型 构象的转变采取齐变方式。 正效应物(如底物)与负效应物浓度的比例决定酶处于何种状态。 由底物调节的效应更符合此模型。
弹性蛋白酶-Ala、Val小的非极性侧链 口诀: 胰蛋白赖精碱 胰凝乳苯色酪 胃蛋白苯色酪缬氨
二、酶促反应机制
酶的催化功能基团能与底物发生瞬间的相互作用, 激活反应的底物。
18
(一)基元催化的分子机制
1. 酸-碱催化 P173
狭义酸-碱催化:H+ 或OH-1 对反应的加速。 广义酸-碱催化:通过质子酸提供部分质子,
别构酶
别构酶的结构特点
1一般是寡聚酶,多个亚基构成,有四级结构; 2.含活性部位和调节部位。 3.调节部位: 酶分子中存在着一些可以与其他
分子发生某种程度的结合的部位,从而引起酶 分子空间构象的变化,对酶起激活或抑制作用。 4.调节部位只能与专一的调节物非共价结合。
43
调节物
调节物:凡能使酶分子发生别构作用的物质。 使酶活性增加的调节物称正调节物,反之为负调节物。
邻近定向效应
2.诱导契合和底物的形变
当酶与底物靠近时,不仅酶构象受底物 作用而变化,底物分子也常常受酶作用而 变化,也就是酶使底物分子中的敏感键发 生“变形”,从而促使底物中的敏感键更 易于破裂。
但是Asp-His-Ser组成的活性位点的排列是相似的。
32
胰蛋白酶、胰凝乳蛋白酶、弹性蛋白 酶的比较
胰蛋白酶:Arg-X, Lys-X 胰凝乳蛋白酶: 芳香族氨基酸 Phe-X, Tyr-X, Trp-X 弹性蛋白酶:Ala-X, Gly-X, Ser-X, Val-X(丙甘撕缬)
原因:酶与底物结合部位,即“口袋”,在结构上的 微小差异引起的。
2、催化基团 催化基团决定酶所催化反应的性质,同时 也是决定反应的高效性。
3、调控基团 对酶起激活或抑制作用的基团。
酶的活性中心示意图
胰蛋白酶活性位点
7
酶活性部位的特点(P172)
1. 只占酶分子总体相当小的部分; 2. 是一个三维实体; 3. 活性部位与底物分子经诱导契合互补; 4. 位于酶分子表面的一个裂缝内; 5. 底物通过较弱的次级键与酶结合; 6. 活性部位具有柔性或可运动性。 7. 酶活性中心的基团主要有:
催化三联体。
His的咪唑基是Asp羧 基和Ser羟基间的桥梁, 形成电荷中继网。
30
胰蛋白酶、胰凝乳蛋白酶、弹性蛋白 酶的比较
结构(一级结构和三维结构)及机理:极其相似
31
催化三联体具有相似的结构
枯草杆菌蛋白酶
胰凝乳蛋白酶
枯草杆菌蛋白酶和胰凝乳蛋白酶结构有很大的不同,而且组成
催化三联体的残基处于一级顺序中不同的位置。
速率也不会发生大的改变。 保证糖酵解的正常进行。
54
五、酶活性共价修饰调节
(一)酶的可逆共价修饰
蛋白质的磷酸化与脱磷酸化过程,是生物体内一 种普遍的调节方式。
由蛋白激酶和蛋白磷酸酶分别催化完成,以ATP 作为磷酸基供体。
底物蛋白质被磷酸化的氨基酸残基主要是Ser、 Thr,个别为Tyr。
可转变成有活性的酶。 酶原中,酶的活性部位被掩盖或者没有形成,酶原
的激活实质上就是酶活性部位形成的过程。
57
内肽酶 胃蛋白酶 胰蛋白酶 胰凝乳蛋白酶 弹性蛋白酶 外肽酶 羧肽酶 氨肽酶
1. 消化系统中酶原激活
消化系统中的酶多以酶原的形式合成和贮存。 酶原在一定条件被激活。
55
磷酸化酶 a(有活性)磷 酸 化酶 磷 酸 酶 磷酸化酶 b(无活性) 磷酸化酶 激 酶
葡萄糖 n Pi 葡萄糖 n1 1 磷酸葡萄糖
56
五、酶活性共价修饰调节 (二)酶原的激活-不可逆共价调节
酶原(无活性的前体酶)(有活性)
酶原:没有活性的酶的前体。 酶原的激活:酶原在一定条件下经适当的物质作用
48
(四)别构酶举例 1. 天冬氨酸转氨甲酰酶(ATCase)
1) 氨甲酰磷酸 2) 两种底物与酶的结合具有正协同效
应,表现为S型的动力学曲线。 正调接物:ATP,作为信号表明有
能量供DNA复制使用并引发需求嘧 啶核苷酸的生物合成 负调节物:CTP,在嘧啶核苷酸已 丰足时,不再进行不必要的氨甲酰 天冬氨酸及其他后续中间产物的合 成
正协同性(同促)相互作用:酶的一个亚基上活性部 位与底物(调节物)结合,增加其余亚基活性部位的亲 和力。 负协同性(异促)相互作用:酶的一个亚基上活性部 位与底物(调节物)结合,降低其余亚基活性部位的亲 和力。
44
(二)别构酶的动力学特征
12 3
P184 图11-16A
1. 无调节物:双曲线 2. 正协同作用: S形曲线 3. 负协同作用:表观双曲线
或是通过质子碱接受部分质子,达到降低反 应活化能的目的的一种作用。 酶参与的一般都是广义的酸-碱催化方式。
19
酶分子中可以作为广义酸、碱的基团
广义酸碱催化:质子供体及质子受体的催化
20
2. 共价催化 P174
催化剂与底物之间瞬间形成共价键以提高反 应速率的一种催化机制。
包括亲核催化和亲电催化。 催化剂通过放出电子或获取电子并作用于底
定酶的专一性;
催化部位 catalytic site:负责催化底物键的断裂 形成新键,决定酶的催化能力和催化反应性质。
3
酶的活性中心是指结合底物和将底物转 化为产物的区域,通常是相隔很远的氨 基酸残基形成的三维实体。
活性中心
结合基团 催化基团 调控基团
中心内必需基团 中心外必需基团
1、结合基团 酶的结合基团决定酶反应的专一性。
P188 图11-21
50
天冬氨酸转氨甲酰酶构象的转变
2个催化3聚体 3个调节2聚体
ATP
CTP P189 图11-22
51
(四)别构酶举例 天冬氨酸转氨甲酰酶(ATCase)
天冬氨酸蛋白酶的结构特点:每一种酶的活 性部位具有两个天冬氨酸残基。
天冬氨酸蛋白酶的作用机制
52
(四)别构酶举例 2. 甘油醛-3-磷酸脱氢酶
的底物(对甲苯磺酰苯丙氨酸乙酯)类似物。
P195: 3
胰蛋白酶的亲和标记试剂
O CH2 CH C CH2Cl
NH SO2 (CH2)4 NH3+ N-(ε -氨基-丁基)磺酰--L-苯丙氨酰氯甲基酮
苯丙氨酸乙酯
丝氨酸蛋白酶底物结合部位:
胰凝乳蛋白酶-芳香环
胰蛋白酶-带正电荷的Lys、Arg进入
胰蛋白酶的口袋底部有一个负电荷的Asp189,能促 进与带正电荷的Arg, Lys结合。
弹性蛋白酶的口袋较浅,只能容纳侧链较小的残基。
33
酶与底物结合部位,即“口袋”,在 结构上有微小差异
P180 图11-12
34
胰凝乳蛋白酶反应的详细机制(1)
底物
结合底物
11
His57 质子供体
23
3-磷酸甘油醛
1,3-二磷酸甘油酸
53
(四)别构酶举例 2. 甘油醛-3-磷酸脱氢酶
负协同效用。有4个亚基,可以和4个[NAD+]结 合。 [NAD+]的结合引起该亚基的构象发生变化, 并降低了同一酶分子中相邻的空位亚基的结合 亲和力。K1<K2<K3<K4。
生理意义: 在[NAD+]浓度很低时,能保证与酶的结合; 在[NAD+]浓度很高时,由于负协同效应,反应
酶
底物 同时形变
产物
底物分子发生形变
三、酶促反应机制的举例
(一)丝氨酸蛋白酶 1. 结构特点: 丝氨酸蛋白酶家族包括: 胰蛋白酶、胰凝乳蛋白酶、弹性蛋白酶、 凝血酶等,前三种酶为消化酶。 丝氨酸残基是酶活性必需的。
29
2. 丝氨酸蛋白酶的作用机制
丝氨酸蛋白酶的三个 极性残基- Ser-195, His-57, and Asp-102
P185 图11-17
P185 图11-18
47
2. KNF模型/序变模型
当配体不存在时,别构酶以T型构象存在,结合配体后由T型向R型 转变;
别构酶以序变方式进行构象变化。 亚基间的相互作用可能是正协同效应,也可能是负协同效应。 此序变模式可以较好的说明非底物调节的效应。
P187 图11-20
23
(二)酶具有高催化能力的原因
1. 底物和酶的邻近效应与定向效应 2. 诱导契合和底物形变 3. 电荷极化和多元催化 4. 疏水的微环境的影响
24
1、 邻近效应、 定向效应
• 邻近效应:在酶促反应中,由于酶和底物分子之间 的亲和性,底物分子有向酶的活性中心靠近的趋势, 使底物在酶活性中心的有效浓度大大增加的效应。
第 十一 章 酶作用机制和酶活性调节
1
主要内容
1. 酶的活性部位及其确定方法(重点难点考点) 2. 酶促反应机制(掌握) 3. 酶促反应机制的举例(重点、考点) 4. 酶活性的别构调节(重点、考点) 5. 酶活性的共价调节(重点) 6. 同工酶(了解)
2
一、酶的活性部位及其确定方法
酶的活性部位:与酶活力直接相关的区域 结合部位 binding site:负责与底物的结合,决
45
(三)协同性配体结合的模型
亚基之间存在协同效应 当一个亚基结合底物或效应物后,该亚基的构
象发生变化,从而影响其他亚基也发生构象变 化。 别构模型:
齐变模型/MWC模型/对称模型 序变模型
46
1. MWC模型/齐变模型/对称模型
每个亚基对一种配体只有一个结合位点; 每种亚基有两种构象, R型 T型 构象的转变采取齐变方式。 正效应物(如底物)与负效应物浓度的比例决定酶处于何种状态。 由底物调节的效应更符合此模型。
弹性蛋白酶-Ala、Val小的非极性侧链 口诀: 胰蛋白赖精碱 胰凝乳苯色酪 胃蛋白苯色酪缬氨
二、酶促反应机制
酶的催化功能基团能与底物发生瞬间的相互作用, 激活反应的底物。
18
(一)基元催化的分子机制
1. 酸-碱催化 P173
狭义酸-碱催化:H+ 或OH-1 对反应的加速。 广义酸-碱催化:通过质子酸提供部分质子,
别构酶
别构酶的结构特点
1一般是寡聚酶,多个亚基构成,有四级结构; 2.含活性部位和调节部位。 3.调节部位: 酶分子中存在着一些可以与其他
分子发生某种程度的结合的部位,从而引起酶 分子空间构象的变化,对酶起激活或抑制作用。 4.调节部位只能与专一的调节物非共价结合。
43
调节物
调节物:凡能使酶分子发生别构作用的物质。 使酶活性增加的调节物称正调节物,反之为负调节物。