核外遗传
合集下载
遗传学经典核外遗传
线粒体是细胞内的代谢中心,1963年发 现线粒体具有DNA。 线粒体具有半自主性,其DNA编码rRNA、 tRNA以及蛋白质 线粒体产生呼吸酶和多种酶系,呼吸酶 的缺乏导致生长迟缓,长成“小菌落”。
线粒体基因组
mtDNA的基因组成 (1)闭合环状DNA (2) 基因数目和排列顺序相同
(3)有D环和2个复制起始点 (4)基因间没有间隔,因此每个基因不可
核密码 终止 Arg Ile Ile Arg 终止 终止 Ile Leu Leu 终止 终止
核糖体的差异
表 20-4 人类和酵母线粒体核糖体和细胞质核糖体的大小
成分
人类(HeLa)
mt
cyto
酵母
mt
Cyto
核糖体
60S
74S
75S
80S
大亚基
45S
60S
53S
60S
小亚基
35S
40S
35S
40S
能都有自己的起动子 (5)某些蛋白质的密码子与核基因通用
密码子不同; (6)mtDNA主要编码rRNA和tRNA分子,氧化
呼吸所需要的酶类少部分亚基。
酵母的小菌落突变
酵母菌可以进行有性生殖,与链孢霉类似。 正常菌落能自发形成部分小菌落(1%2%),化学诱变剂能诱导小菌落。 小菌落:细胞生长缓慢,长成很小的菌落。 缺少细胞色素a、b以及细胞色素c氧化酶。 稳定遗传。
线粒体中密码子的改变
生物 共同(人、果蝇、酵母) 哺乳动物 哺乳动物 果蝇 果蝇 纤毛虫 另一类纤毛虫 酵母 酵母 酵母 酵母 支原体
密码子 UGA AGA/AGG AUA AUA AGA UAA/UAG UGA/UAG AUA CCA CUG UAG UGA
线粒体基因组
mtDNA的基因组成 (1)闭合环状DNA (2) 基因数目和排列顺序相同
(3)有D环和2个复制起始点 (4)基因间没有间隔,因此每个基因不可
核密码 终止 Arg Ile Ile Arg 终止 终止 Ile Leu Leu 终止 终止
核糖体的差异
表 20-4 人类和酵母线粒体核糖体和细胞质核糖体的大小
成分
人类(HeLa)
mt
cyto
酵母
mt
Cyto
核糖体
60S
74S
75S
80S
大亚基
45S
60S
53S
60S
小亚基
35S
40S
35S
40S
能都有自己的起动子 (5)某些蛋白质的密码子与核基因通用
密码子不同; (6)mtDNA主要编码rRNA和tRNA分子,氧化
呼吸所需要的酶类少部分亚基。
酵母的小菌落突变
酵母菌可以进行有性生殖,与链孢霉类似。 正常菌落能自发形成部分小菌落(1%2%),化学诱变剂能诱导小菌落。 小菌落:细胞生长缓慢,长成很小的菌落。 缺少细胞色素a、b以及细胞色素c氧化酶。 稳定遗传。
线粒体中密码子的改变
生物 共同(人、果蝇、酵母) 哺乳动物 哺乳动物 果蝇 果蝇 纤毛虫 另一类纤毛虫 酵母 酵母 酵母 酵母 支原体
密码子 UGA AGA/AGG AUA AUA AGA UAA/UAG UGA/UAG AUA CCA CUG UAG UGA
遗传学第八章 核外遗传分析课件

遗传学 第八章 核外遗传分析
2、 性比(sex-ratio,SR)因子
◇SR因子是胞质中的一种原生动物,在雌蝇 和雄蝇中都能发现,但对发育中的雄性幼 虫是致死的,所以后代中雌蝇比例远大于 雄蝇比例。
◇将SR雌蝇的卵细胞质注入正常雌蝇可诱导 SR现象。
◇有证据表明,产生雄性致死毒素的可能是原 生动物内的病毒。
基因型与表型的关系
细胞质基因 正常(N) 不育(S)
核基因型
RfRf(可育) Rfrf(可育) rfrf(不育) N(RfRf)可育 N(Rfrf)可育 N(rfrf)可育 S(RfRf)可育 S(Rfrf)可育 S(rfrf)不育
遗传学 第八章 核外遗传分析
(二)可能的遗传机制 1、线粒体与雄性不育的关系 2、叶绿体与雄性不育的关系
遗传学 第八章 核外遗传分析
遗传学 第八章 核外遗传分析
• KSS综合征(Keams-Sayre Syndrome)是多 系统线粒体病,主要症状为眼肌麻痹和色素性视 网膜炎。患者骨骼肌细胞mtDNA有2.0kb7.0kb的缺失。 遗传学 第八章 核外遗传分析
五、叶绿体遗传及其分子基础
(一)衣藻的叶绿体遗传
◇ 衣藻细胞中只有一个叶绿体,约含50个 拷贝的环状ds-DNA分子。 ◇ 不同交配型(mt+,mt-)的单倍体衣藻 杂交,形成短暂2n时期,进行减数分裂。 ◇ 虽然杂交双方融合时为合子提供等量细 胞质,但叶绿体只由mt+方传递,表现单亲 遗传。
遗传学 第八章 核外遗传分析
遗传学 第八章 核外遗传分析
(二)叶绿体遗传的分子基础 1、叶绿体基因组
大小:环状双链DNA分子。大小120-190kb。 其基因序列中不含5-甲基胞嘧啶。
◆ cpDNA编码约100种蛋白质和RNAs,包 括45个编码RNA的基因,27个编码与基因表达 有关的蛋白的基因,18个编码类囊体膜的蛋白基 因和10个与电子传递功能有关的基因。
2、 性比(sex-ratio,SR)因子
◇SR因子是胞质中的一种原生动物,在雌蝇 和雄蝇中都能发现,但对发育中的雄性幼 虫是致死的,所以后代中雌蝇比例远大于 雄蝇比例。
◇将SR雌蝇的卵细胞质注入正常雌蝇可诱导 SR现象。
◇有证据表明,产生雄性致死毒素的可能是原 生动物内的病毒。
基因型与表型的关系
细胞质基因 正常(N) 不育(S)
核基因型
RfRf(可育) Rfrf(可育) rfrf(不育) N(RfRf)可育 N(Rfrf)可育 N(rfrf)可育 S(RfRf)可育 S(Rfrf)可育 S(rfrf)不育
遗传学 第八章 核外遗传分析
(二)可能的遗传机制 1、线粒体与雄性不育的关系 2、叶绿体与雄性不育的关系
遗传学 第八章 核外遗传分析
遗传学 第八章 核外遗传分析
• KSS综合征(Keams-Sayre Syndrome)是多 系统线粒体病,主要症状为眼肌麻痹和色素性视 网膜炎。患者骨骼肌细胞mtDNA有2.0kb7.0kb的缺失。 遗传学 第八章 核外遗传分析
五、叶绿体遗传及其分子基础
(一)衣藻的叶绿体遗传
◇ 衣藻细胞中只有一个叶绿体,约含50个 拷贝的环状ds-DNA分子。 ◇ 不同交配型(mt+,mt-)的单倍体衣藻 杂交,形成短暂2n时期,进行减数分裂。 ◇ 虽然杂交双方融合时为合子提供等量细 胞质,但叶绿体只由mt+方传递,表现单亲 遗传。
遗传学 第八章 核外遗传分析
遗传学 第八章 核外遗传分析
(二)叶绿体遗传的分子基础 1、叶绿体基因组
大小:环状双链DNA分子。大小120-190kb。 其基因序列中不含5-甲基胞嘧啶。
◆ cpDNA编码约100种蛋白质和RNAs,包 括45个编码RNA的基因,27个编码与基因表达 有关的蛋白的基因,18个编码类囊体膜的蛋白基 因和10个与电子传递功能有关的基因。
《核外遗传分析》课件

核外遗传的特点
核外基因组结构简单,基 因密度高,复制和转录过 程相对简单。
核外基因组具有母系遗传 的特点,即线粒体基因组 和叶绿体基因组只来自母 本。
核外基因组在进化上相对 保守,但在某些物种中也 可能发生基因重组和突变 。
核外遗传分析的意义
核外遗传分析有助于深入了解生 物的进化历程和物种起源。
详细描述
表观遗传学分析在核外遗传分析中扮演着重要的角色。表观遗传学标记如DNA甲基化和 组蛋白修饰等可以影响基因的表达水平,进而影响生物体的表型。通过分析这些表观遗 传学标记,可以深入了解基因表达的调控机制,为疾病诊断和治疗提供新的思路和方法
。
转录组学分析
总结词
转录组学分析主要研究基因转录本的种类和丰度,通过分析转录本的变化,揭示基因表达的动态变化 和调控机制。
和思路。
02
个体化用药
根据个体的核外遗传变异情况,可以制定针对个体的个性化用药方案,
提高药物的疗效并降低副作用。
03
药物反应差异研究
不同个体对同一种药物的反应可能存在差异,这种差异可能与个体的核
外遗传变异有关,通过研究这种关系可以更好地理解药物反应的个体差
异。
个体化医疗的核外遗传分析应用
精准医疗
基于个体的核外遗传变异情况,可以为患者提供精准、个性化的 治疗方案,提高治疗效果。
技术发展的挑战
技术更新换代
01
核外遗传分析技术不断发展,需要不断更新知识和技术,以适
应新的分析需求。
技术局限性
02
目前核外遗传分析技术仍存在一定的局限性,如检测灵敏度、
特异性等方面仍有待提高。
技术普及
03
如何将核外遗传分析技术普及到临床实践中,提高其在医学研
第十章核外遗传分析

核基因 A:a=1:1,表型全部为poky突变
44
1.4 粗糙脉孢菌 poky 小菌落遗传
野生型与 poky 突变体旳菌丝融合 →异核体 →异核体内两种不同核形成两种不同旳分生 孢子→不同旳菌株: 有野生型核旳菌株体现出 poky 小菌落性状, 有 poky 小菌落性状旳菌株却体现出野生型菌 落旳性状.
42
1.3 链孢霉缓慢生长突变型(Poky)旳遗传
正交:
正常 mt A
(+) (野生型)
缺失 mt
a
接合时只提供核
(-) (poky)
A AAAa a a a
子囊孢子从“+” 接合型那里得到正常线粒体,全部野生型。 核基因 A:a=1:1
43
反交:
A
(+) (poky)
a
(-) (野生型)
A AAA a a a a
34
第四节 线粒体遗传及其遗传基础
35
1. 线粒体遗传
1.1 酵母旳小菌落遗传
野生型酵母生长快, 形成旳菌落 较 大, 所以称大菌落( grande) 20 世纪40 年代,法国学者埃弗雷西(B. Ephrassi)在面包 酵母中发觉了一种突变型生长缓慢, 形成旳菌落很小, 所 以称小菌落突变体(petite mutant)。 后来发觉该突变体缺乏 细胞色素 a, b 和细胞色素c氧化酶, 是线立体基因全部丢失(突变)旳成果。
8
5. 有细胞质中旳附加体或共生微生物决定旳性状, 其体现类似病毒旳转导或感染,即可传递给其他 细胞。
6. 基因定位困难。 因为1)杂交后裔不体既有百分 比旳分离;2)带有保值基因旳细胞器在细胞分裂 时旳分配不是均匀旳。
9
核外遗传现象旳发觉
44
1.4 粗糙脉孢菌 poky 小菌落遗传
野生型与 poky 突变体旳菌丝融合 →异核体 →异核体内两种不同核形成两种不同旳分生 孢子→不同旳菌株: 有野生型核旳菌株体现出 poky 小菌落性状, 有 poky 小菌落性状旳菌株却体现出野生型菌 落旳性状.
42
1.3 链孢霉缓慢生长突变型(Poky)旳遗传
正交:
正常 mt A
(+) (野生型)
缺失 mt
a
接合时只提供核
(-) (poky)
A AAAa a a a
子囊孢子从“+” 接合型那里得到正常线粒体,全部野生型。 核基因 A:a=1:1
43
反交:
A
(+) (poky)
a
(-) (野生型)
A AAA a a a a
34
第四节 线粒体遗传及其遗传基础
35
1. 线粒体遗传
1.1 酵母旳小菌落遗传
野生型酵母生长快, 形成旳菌落 较 大, 所以称大菌落( grande) 20 世纪40 年代,法国学者埃弗雷西(B. Ephrassi)在面包 酵母中发觉了一种突变型生长缓慢, 形成旳菌落很小, 所 以称小菌落突变体(petite mutant)。 后来发觉该突变体缺乏 细胞色素 a, b 和细胞色素c氧化酶, 是线立体基因全部丢失(突变)旳成果。
8
5. 有细胞质中旳附加体或共生微生物决定旳性状, 其体现类似病毒旳转导或感染,即可传递给其他 细胞。
6. 基因定位困难。 因为1)杂交后裔不体既有百分 比旳分离;2)带有保值基因旳细胞器在细胞分裂 时旳分配不是均匀旳。
9
核外遗传现象旳发觉
第十三章-核外遗传分析

裂时分配是不均匀的。
细胞质遗传:正交和反交的遗传表现不同。
核遗传:表现相同,其状表现于母本时才能遗传给子代,
故又称母性遗传。
母性遗传:
真核生物有性过程:
卵细胞:有细胞核、大量的细胞质和细胞器。
∴能为子代提供核基因和全部或绝大部分胞质基因。
精细胞:只有细胞核,细胞质或细胞器极少或没有。
mtDNA被恢复。在校正事情中有时出现mtDNA的重排,因为
在线粒体基因组中编码蛋白质的基因是广泛散布的,这些缺失
和重排导致有氧呼吸中酶的缺陷,而产生小菌落。
人们提出抑制小菌落对正常的线粒体有抑制效应可能是:
(1)、抑制型线粒体mtDNA可能比正常的线粒体复制得更快,
在细胞中简单地蔓延,所观察的在品种间的抑制性的变异,
DNA进行了比较研究,发现小菌落线粒体DNA除有一些片断
多次重复外,线粒体基因组中还有大片段缺失。可见酵母小菌
落突变是由于线粒体DNA的遗传变异,致使线粒体不能正常执
行其功能,线粒体中蛋白质合成受阻,造成了呼吸代谢的缺陷。
3、抑制小菌落(suppressive petites)
抑制小菌落对野生型产生影
倍体细胞融合产生二倍体对细胞质是大小相同贡献相等的。
中性小菌落的本质是什么?
根据氯化铯密度梯度离心,测出小菌落细胞中线粒体DNA
与大菌落的线粒体DNA明显不同,有时小菌落中甚至还测不出
线粒体DNA 的存在。这说明,小菌落突变使线粒体DNA严重
缺损或大部分丢失。用分子杂交和限制酶分析方法对小菌落线
粒体DNA全部缺失或严重变异的类型与野生型大菌落的线粒体
S至80 S大小不等的颗粒,由两个大小不等的亚基组成,每个亚基只
有一条由线粒体DNA转录而来的rRNA分子。
细胞质遗传:正交和反交的遗传表现不同。
核遗传:表现相同,其状表现于母本时才能遗传给子代,
故又称母性遗传。
母性遗传:
真核生物有性过程:
卵细胞:有细胞核、大量的细胞质和细胞器。
∴能为子代提供核基因和全部或绝大部分胞质基因。
精细胞:只有细胞核,细胞质或细胞器极少或没有。
mtDNA被恢复。在校正事情中有时出现mtDNA的重排,因为
在线粒体基因组中编码蛋白质的基因是广泛散布的,这些缺失
和重排导致有氧呼吸中酶的缺陷,而产生小菌落。
人们提出抑制小菌落对正常的线粒体有抑制效应可能是:
(1)、抑制型线粒体mtDNA可能比正常的线粒体复制得更快,
在细胞中简单地蔓延,所观察的在品种间的抑制性的变异,
DNA进行了比较研究,发现小菌落线粒体DNA除有一些片断
多次重复外,线粒体基因组中还有大片段缺失。可见酵母小菌
落突变是由于线粒体DNA的遗传变异,致使线粒体不能正常执
行其功能,线粒体中蛋白质合成受阻,造成了呼吸代谢的缺陷。
3、抑制小菌落(suppressive petites)
抑制小菌落对野生型产生影
倍体细胞融合产生二倍体对细胞质是大小相同贡献相等的。
中性小菌落的本质是什么?
根据氯化铯密度梯度离心,测出小菌落细胞中线粒体DNA
与大菌落的线粒体DNA明显不同,有时小菌落中甚至还测不出
线粒体DNA 的存在。这说明,小菌落突变使线粒体DNA严重
缺损或大部分丢失。用分子杂交和限制酶分析方法对小菌落线
粒体DNA全部缺失或严重变异的类型与野生型大菌落的线粒体
S至80 S大小不等的颗粒,由两个大小不等的亚基组成,每个亚基只
有一条由线粒体DNA转录而来的rRNA分子。
遗传学(第3版)第5章核外遗传分析

sm-r mt + sm-s mt - 全部后代(99%)表现为sm-r sm-s mt + sm-r mt - 全部后代(99%)表现为sm-s
以上结果表明正反交结果不同,说明衣藻对链霉素的抗性或 敏感是由非孟德尔遗传因子所控制(图5-3)
研究表明:该性状由衣藻中叶绿体基
因组DNA上的基因所控制。交配型mt+ 和mt–细胞质中都含有叶绿体基因组 DNA,当发生交配时,两个亲本对合 子贡献的细胞质是相等的,而后代总 是保留mt+亲本与叶绿体基因组上控 制的性状有关的表型。一种分子解释
某些产物积累在卵母细胞的细胞质中,使子代表型不由自身 的基因型所决定而出现与母体表型相同的遗传现象,则称为 母体影响。 母体影响有两种: 一种是短暂的,只影响子代个体的幼龄期;
5.1.3衣藻抗生素抗性的遗传
衣藻通常行无性生殖,有时通过两种形态相同但交配型
不同的配子进行融合,行有性生殖。同型配子间不能融合,
仅相反交配型的配子能够融合产生二倍体的合子。衣藻的交 配型是由细胞核内一对等位基因mt +和mt -所决定的。配子
融合给合子提供了相等的细胞内含物,合子萌发时,通常立
即发生减数分裂,四个减数分裂的产物作为非顺序四分子在 一个子囊中,其中有两个mt +细胞和两个mt -细胞,即核基
②自体受精(Autogamy): 同一个体的两个小核经减数分裂,留下一个小核, 该小核分裂一次又相互合并,随后再分裂发育成小核和大核。 (Fig--45 ) 3).草履虫放毒型和敏感型的接合 若 某些草履虫 草履虫素 杀死其它的草履虫但对自己无害。 能产生草履虫素的个体叫做放毒型 而受害的个体就称为敏感型 根据遗传学实验,草履虫的放毒型必需有两种因子同时存在: ① 细胞质因子,叫做卡巴粒(Kappa particle , κ)
核外遗传

生可育的花粉,但能产生正常的卵细胞,可
接受外来花粉而受精结籽。
产生原因
环境因素:如高温杀雄(不遗传) 遗传因素:可遗传
(一)植物雄性不育的类型
Types of male sterility
根据雄性不育发生的遗传机制,Sears(1947)将 之分为三种类型。 1. 细胞质雄性不育(cytoplasmic male sterile CMS)
(4)某些蛋白质的密码子与核基因通用密码子不同。
Cytb D-loop ND6 ND5 12S rRNA 16S rRNA ND4 ND4L ND3 CO3 ATPase6 ATPase8 CO2 CO1 TRNA genes; Coding regions, Indicates direction of gene , 5to3’ CO: cytochrome oxidase; ND: NADH dehydrogenase Gig. Human mitochondrial DNA has 22 tRNA genes,2 rRNA genes,ahd 13 protein-coding regions.14 of the 15 protein-coding or rRNA-coding regions are transcribed in the same direction.14 of the tRNA genes are expressed in the clockwise direction and 8 are read countr clockwise. ND2 ND1
能产生草履虫素的品系——放毒型; 不产生草履虫素的品系——敏感型 草履虫的放毒型既与细胞质有关,也与细 胞核有关。 放毒型为:K_ + κ
敏感型为:kk
第八章 核外遗传

外包双层膜、直径为0.2~0.4μm的小颗粒,含DNA、
RNA和酶。
卡巴粒的rRNA能与E. coli DNA杂交,但不能与
草履虫的DNA杂交。
认为卡巴粒是一种共生型的具有螺旋结构的放毒
型细菌。
其它放毒性颗粒:γ粒、λ粒、μ粒。
二、果蝇的感染性遗传
果蝇对CO2的敏感性的遗传
果蝇“性比(SR)失常”
突变型a
不产生犬尿氨酸,幼虫皮肤无色,成 虫复眼红色。
(能合成犬尿氨酸)
麦粉蛾的色素遗传中的母体影响
无色 结果同一般测交
是短暂的,只影响子代的幼龄期。
2、持久的母体影响
(d)
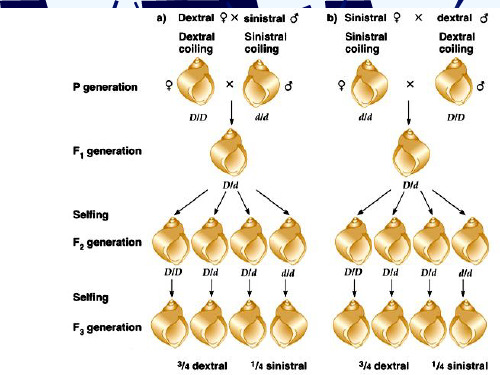
椎实螺的外壳
(D)
椎实螺外壳旋向的遗传
母体影响
母体 影响 母体中核基因D的产物积累在卵母细胞的 细胞质中,使F2中的dd个体也表现右旋.
线粒体,小菌落消失。
二、线粒体基因组
线粒体基因组的一般性质
线粒体DNA的组成和蛋白
质合成
人类的线粒体基因病
(一) 线粒体基因组的一般性质
1962年,由KIS等发现线 粒 体 ( mitochondrion ) 内有DNA(mtDNA), 与核内DNA明显不同
环状(线状)、裸露
不同生物线粒体DNA的比较
酵母菌
固体培养基上通气培养
大菌落
缺少细胞色 素a,b和细 小菌落 胞色素c氧 1-2% 化酶—呼吸 缺陷突变株
大菌落
小菌落 1-2%
小菌落
都为正常型菌落
解 释
小菌落的形成:线粒体DNA的遗传变
异。
小菌落与正常的酵母杂交:细胞质融
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
ቤተ መጻሕፍቲ ባይዱ
些细胞质基因在遗传时并不表现母系遗传现象)
2. 遗传方式是非孟德尔式的,杂交后代一般不出 现一定的分离比例。 3. 连续的回交能把母本的核基因全部置换掉,但 母本的细胞质基因及其控制的性状仍不消失
第二节 叶绿体遗传
1. 叶绿体的遗传表现
紫茉莉的花斑叶色遗传
天竺葵的花斑叶色遗传
玉米叶片的埃型条纹的遗传
2、
3、 4、 5、
S(RR)1
保持系 ——N(rr) 5
质核不育型的三系配套(三系二区)
S rr
不育系
×
N rr 保持系
S rr
×
N RR
恢复系
S rr
S Rr F1杂交种 N rr N RR
光温敏雄性不育性与两系法
光温敏雄性不育性:
不育性表现受遗传与环境因素共同影响,也称 为生态不育型
从实验结果看出:
正反交结果不同,子一代总是表现出母本的性状 ,
与父本提供的花粉无关。
原因:精卵结合中形成的合子父母双亲所提供的 遗传物质不均等造成的。
细胞质遗传的主要特点
核外遗传因子由一个亲本而来的 ,不经过有丝分裂
或减数分裂,它们的行为不按核基因的方式进行:
1. 正反交的结果不一样,F1表现母系遗传。(有
减数分裂产物时线粒体随机分配结果:
全部为野生型菌落(+/-)
或混合型菌落
• 野生型菌落(+/-, +) • 小菌落(-) 抑制型小菌落
中性小菌落的遗传
由于缺乏线粒体基因组,故 完全不具有线粒体负责的
ATP合成电子传递途径(有
氧呼吸)
细胞生长的能量来源依赖于
细胞质基质中进行的发酵途 径(无氧呼吸/糖酵解)
第四节 草履虫的放毒性遗传 (共生体遗传)
某些真核生物细胞质中,除质体、线粒体等必
需的成分外,还存在一些非必需的有形成分,它们
以某种共生的形式存在于细胞中,称为共生体。 典型的共生体遗传实例是草履虫卡巴粒 (Kappa particles,k)的遗传。
草履虫的细胞结构及生殖方式
细胞结构:草履虫含有两种细胞核—大核和小核。
10%的结构与功能蛋白;其余部分由核基因编 码
线粒体遗传的分子基础
线粒体DNA(mtDNA)的分子形态:
闭合环状双链分子
拷贝数:多拷贝
功能:mtDNA编码少部分线粒体结构与功能蛋白、RNA 遗传体系:
具有与核DNA相对独立的复制、转录和翻译系统 甚至具有一些与核基因组不同的遗传密码
两者必须同时存在才能保证放毒型的稳定性。
K/K 或 K/k
+ 卡巴粒
放毒型
k/k, K/K 或 K/k
敏感型
纯合放毒型(KK+卡巴粒)与敏感型(kk)杂交 短时接合:两亲本只交换了小核而未交换细胞质 接合时间长:两亲本不仅交换了小核,而且还 交换了部分细胞质。
短时接合
放毒型 敏感型 K
K 放毒型 敏感型
3. 子代表型出现一定的分离比。
某研究所依据“多莉羊”的克隆技术路线克 隆了一头羊,研究者发现该克隆羊具有一些
提供卵子个体的相似表型,请分析这种情况
出现的原因。(至少举出两种原因)
l ; l 并且卵子中有供体母性效应基因的表达产物, 因 由于提供卵子的供体提供了全部的细胞质DNA
此克隆体会出现部分类似卵子供体的表型
在包括玉米、小麦在内的80多种高等植物中发现。
细胞质雄性不育性可以得到保持(繁殖不育系)
以不育系作为母本、以可育品系作为父本,后代总是
具有不育胞质,因而也是不育系
但是不能够得到恢复
不能够生产杂交种
3. 核质互作不育型
细胞质不育基因S和核内隐性基因rr共同作用的结果
细胞质遗传类型
雄性不育的利用
雄性不育的概念
不育(sterility):个体不能产生有功能的配子或 不能产生在一定条件下能够存活的合子的现象。
雄性不育(male sterility):植株不能产生正常
的花药、花粉或雄配子的不育类型。
但其雌蕊发育正常,能接受正常
花粉而受精结实。
植物雄性不育的类型
1. 核不育型:
2. 叶绿体的遗传特点
天竺葵的花斑叶色遗传
母本枝条颜色 父本枝条颜色 后代表型
大部分绿色 绿色 花 斑 少部分花斑 大部分花斑 花斑 绿 色
少部分绿色
玉米叶片的埃型条纹的遗传
玉米第7染色体上(核基因) 正常绿色叶(Ij);
白色,条纹叶(ij)
条纹叶基因纯合
(ijij)时,会引起 叶绿体基因变异;
多数核不育型均受简单的一对隐性基因(ms)控制, 纯合子(msms)表现雄性不育。
雄性不育性的恢复; 雄性不育性的保持 msms×MsMs msms×Msms ↓ ↓ Msms (生产杂交种) Msms msms (繁殖不育系)
用普通遗传学的方法不能使整个群体保持其不育性
2. 细胞质不育型
由细胞质基因决定的雄性不育类型。此类型已
poky突变体与野生型进行正、反交
poky ♀×♂野生型 野生型♀×♂poky 野生型 野生型
poky突变型 poky突变型
解释:线粒体上与细胞色素氧化酶有关的基因突变
缓慢生长突变型在幼嫩培养阶段时无细胞色素氧化酶, 观察到缓慢生长突变型中的线粒体结构不正常。
啤酒酵母的小菌落突变
菌落大小性状(生长速度)遗传
可育花粉均为带显性可育基因R的花粉 受花粉粒基因型控制,败育发生较晚
不育性难以保持,在杂种优势利用中无利用价值
第六节 母性影响 (maternal
effect)
母体基因型决定后代表型的现象称为母性影响
1. 持久的母性影响 2. 短暂的母性影响
1. 持久的母性影响
例如:椎实螺外壳旋转方向 ♂ dd ♀ DD × 左旋 右旋 Dd 右旋
1 DD 右旋 右旋
2 Dd 右旋 右旋
1 dd 右旋
左旋
♀ dd ♂DD 左旋 × 右旋 Dd 左旋 1 DD 右旋 右旋 2 Dd 右旋 右旋 1 dd 右旋 左旋
持久性母性影响的遗传机理
椎实螺外壳的旋转方向是由受精卵第一次和第二次卵裂时
纺垂体分裂的方向决定的。纺锤体的方向决定于卵细胞的特性, 卵细胞的特性又决定于母体的基因型。
第三节 线粒体遗传
1. 线粒体的遗传表现
红色面包霉的生长缓慢突变
啤酒酵母的小菌落突变 2. 线粒体遗传的特点
红色面包霉的生长缓慢突变
红色面包霉的两种有性生殖方式: 菌丝体融合 分生孢子(相当于精子)与原子囊果(相当于卵细胞) 融合 红色面包霉有一种生长缓慢突变(poky)
遗传类型能够通过无性生殖稳定遗传 正反交的遗传表现不同
叶绿体DNA (ctDNA) 的分子形态: 闭合环状双链分子 拷贝数:多拷贝 功能: ctDNA编码部分叶绿体结构与功能蛋白、RNA 其它叶绿体结构与功能蛋白由核基因编码 遗传体系: 具有与核DNA相对独立的复制、转录和翻译系统
核糖体:70S(50S+30S),其23S、4.5S、5S、16SrRNA 基因以及1/3核糖体蛋白质都由叶绿体DNA编码。 30多个tRNA基因都由叶绿体DNA编码
且以细胞质遗传
的方式稳定遗传
2. 叶绿体的遗传特点
叶绿体具有遗传自主性: 既能自我复制,又能独立表达(决定性状表现)
叶绿体遗传的自主性是相对的:
在某些情况下受核基因影响(甚至产生细胞质 基因突变)
*叶绿体遗传的分子基础
1962年,由Kis和Plant首先明确肯定植物细胞的 叶绿体中含有DNA。
N:具有正常的细胞质 (可育胞质)
S:具有不育的细胞质 (不育胞质)
RR:显性纯合可育核基因型 Rr:杂合可育核基因型 rr:隐性纯合不育核基因型
细胞核的遗传类型:可育(R) 对不育(r)为显性
不育系
以不育系作为母本,5种可育类型作为父本,后代
的育性遗传表现如图:
1、
不育系——S(rr) 恢复系——N(RR)4
核外遗传
第一节 核外遗传的概念和特点
核外遗传:
由位于核或类核体以外的遗传物质所
决定的遗传现象和遗传规律,也称为细胞 质遗传,非孟德尔遗传.
细胞质遗传的实例和特点
紫茉莉的花斑遗传 1909年由Carl Corrans 发现(irabilis jalapa)
紫茉莉花斑植物杂交的结果 母本枝条表型 白色 父本枝条表型 白色 绿色 花斑 绿色 白色 绿色 花斑 花斑 白色 绿色 花斑 后代表型 白色 白色 白色 绿色 绿色 绿色 花斑,绿色,白色 花斑,绿色,白色 花斑,绿色,白色
人类线粒体疾病
典型的母系遗传,
只有女性才能传递
线粒体疾病
同一家系中不同患
者表现症状有差 异—(异质性)
2. 线粒体遗传的特点
与质体的遗传相比,线粒体基因组的遗传与核基 因组间的关系更紧密
一方面,线粒体在遗传具有半自主性,能够自
我复制、转录和翻译表达 (决定性状表现)
另一方面,线粒体基因组编码线粒体中不到
这种由母体基因型决定子代性状表现的母性影响又叫前定
作用(per-determination)、或延迟遗传(delayed inheritance)。
螺类卵裂的方式示意图
1、2为第一次卵裂,
些细胞质基因在遗传时并不表现母系遗传现象)
2. 遗传方式是非孟德尔式的,杂交后代一般不出 现一定的分离比例。 3. 连续的回交能把母本的核基因全部置换掉,但 母本的细胞质基因及其控制的性状仍不消失
第二节 叶绿体遗传
1. 叶绿体的遗传表现
紫茉莉的花斑叶色遗传
天竺葵的花斑叶色遗传
玉米叶片的埃型条纹的遗传
2、
3、 4、 5、
S(RR)1
保持系 ——N(rr) 5
质核不育型的三系配套(三系二区)
S rr
不育系
×
N rr 保持系
S rr
×
N RR
恢复系
S rr
S Rr F1杂交种 N rr N RR
光温敏雄性不育性与两系法
光温敏雄性不育性:
不育性表现受遗传与环境因素共同影响,也称 为生态不育型
从实验结果看出:
正反交结果不同,子一代总是表现出母本的性状 ,
与父本提供的花粉无关。
原因:精卵结合中形成的合子父母双亲所提供的 遗传物质不均等造成的。
细胞质遗传的主要特点
核外遗传因子由一个亲本而来的 ,不经过有丝分裂
或减数分裂,它们的行为不按核基因的方式进行:
1. 正反交的结果不一样,F1表现母系遗传。(有
减数分裂产物时线粒体随机分配结果:
全部为野生型菌落(+/-)
或混合型菌落
• 野生型菌落(+/-, +) • 小菌落(-) 抑制型小菌落
中性小菌落的遗传
由于缺乏线粒体基因组,故 完全不具有线粒体负责的
ATP合成电子传递途径(有
氧呼吸)
细胞生长的能量来源依赖于
细胞质基质中进行的发酵途 径(无氧呼吸/糖酵解)
第四节 草履虫的放毒性遗传 (共生体遗传)
某些真核生物细胞质中,除质体、线粒体等必
需的成分外,还存在一些非必需的有形成分,它们
以某种共生的形式存在于细胞中,称为共生体。 典型的共生体遗传实例是草履虫卡巴粒 (Kappa particles,k)的遗传。
草履虫的细胞结构及生殖方式
细胞结构:草履虫含有两种细胞核—大核和小核。
10%的结构与功能蛋白;其余部分由核基因编 码
线粒体遗传的分子基础
线粒体DNA(mtDNA)的分子形态:
闭合环状双链分子
拷贝数:多拷贝
功能:mtDNA编码少部分线粒体结构与功能蛋白、RNA 遗传体系:
具有与核DNA相对独立的复制、转录和翻译系统 甚至具有一些与核基因组不同的遗传密码
两者必须同时存在才能保证放毒型的稳定性。
K/K 或 K/k
+ 卡巴粒
放毒型
k/k, K/K 或 K/k
敏感型
纯合放毒型(KK+卡巴粒)与敏感型(kk)杂交 短时接合:两亲本只交换了小核而未交换细胞质 接合时间长:两亲本不仅交换了小核,而且还 交换了部分细胞质。
短时接合
放毒型 敏感型 K
K 放毒型 敏感型
3. 子代表型出现一定的分离比。
某研究所依据“多莉羊”的克隆技术路线克 隆了一头羊,研究者发现该克隆羊具有一些
提供卵子个体的相似表型,请分析这种情况
出现的原因。(至少举出两种原因)
l ; l 并且卵子中有供体母性效应基因的表达产物, 因 由于提供卵子的供体提供了全部的细胞质DNA
此克隆体会出现部分类似卵子供体的表型
在包括玉米、小麦在内的80多种高等植物中发现。
细胞质雄性不育性可以得到保持(繁殖不育系)
以不育系作为母本、以可育品系作为父本,后代总是
具有不育胞质,因而也是不育系
但是不能够得到恢复
不能够生产杂交种
3. 核质互作不育型
细胞质不育基因S和核内隐性基因rr共同作用的结果
细胞质遗传类型
雄性不育的利用
雄性不育的概念
不育(sterility):个体不能产生有功能的配子或 不能产生在一定条件下能够存活的合子的现象。
雄性不育(male sterility):植株不能产生正常
的花药、花粉或雄配子的不育类型。
但其雌蕊发育正常,能接受正常
花粉而受精结实。
植物雄性不育的类型
1. 核不育型:
2. 叶绿体的遗传特点
天竺葵的花斑叶色遗传
母本枝条颜色 父本枝条颜色 后代表型
大部分绿色 绿色 花 斑 少部分花斑 大部分花斑 花斑 绿 色
少部分绿色
玉米叶片的埃型条纹的遗传
玉米第7染色体上(核基因) 正常绿色叶(Ij);
白色,条纹叶(ij)
条纹叶基因纯合
(ijij)时,会引起 叶绿体基因变异;
多数核不育型均受简单的一对隐性基因(ms)控制, 纯合子(msms)表现雄性不育。
雄性不育性的恢复; 雄性不育性的保持 msms×MsMs msms×Msms ↓ ↓ Msms (生产杂交种) Msms msms (繁殖不育系)
用普通遗传学的方法不能使整个群体保持其不育性
2. 细胞质不育型
由细胞质基因决定的雄性不育类型。此类型已
poky突变体与野生型进行正、反交
poky ♀×♂野生型 野生型♀×♂poky 野生型 野生型
poky突变型 poky突变型
解释:线粒体上与细胞色素氧化酶有关的基因突变
缓慢生长突变型在幼嫩培养阶段时无细胞色素氧化酶, 观察到缓慢生长突变型中的线粒体结构不正常。
啤酒酵母的小菌落突变
菌落大小性状(生长速度)遗传
可育花粉均为带显性可育基因R的花粉 受花粉粒基因型控制,败育发生较晚
不育性难以保持,在杂种优势利用中无利用价值
第六节 母性影响 (maternal
effect)
母体基因型决定后代表型的现象称为母性影响
1. 持久的母性影响 2. 短暂的母性影响
1. 持久的母性影响
例如:椎实螺外壳旋转方向 ♂ dd ♀ DD × 左旋 右旋 Dd 右旋
1 DD 右旋 右旋
2 Dd 右旋 右旋
1 dd 右旋
左旋
♀ dd ♂DD 左旋 × 右旋 Dd 左旋 1 DD 右旋 右旋 2 Dd 右旋 右旋 1 dd 右旋 左旋
持久性母性影响的遗传机理
椎实螺外壳的旋转方向是由受精卵第一次和第二次卵裂时
纺垂体分裂的方向决定的。纺锤体的方向决定于卵细胞的特性, 卵细胞的特性又决定于母体的基因型。
第三节 线粒体遗传
1. 线粒体的遗传表现
红色面包霉的生长缓慢突变
啤酒酵母的小菌落突变 2. 线粒体遗传的特点
红色面包霉的生长缓慢突变
红色面包霉的两种有性生殖方式: 菌丝体融合 分生孢子(相当于精子)与原子囊果(相当于卵细胞) 融合 红色面包霉有一种生长缓慢突变(poky)
遗传类型能够通过无性生殖稳定遗传 正反交的遗传表现不同
叶绿体DNA (ctDNA) 的分子形态: 闭合环状双链分子 拷贝数:多拷贝 功能: ctDNA编码部分叶绿体结构与功能蛋白、RNA 其它叶绿体结构与功能蛋白由核基因编码 遗传体系: 具有与核DNA相对独立的复制、转录和翻译系统
核糖体:70S(50S+30S),其23S、4.5S、5S、16SrRNA 基因以及1/3核糖体蛋白质都由叶绿体DNA编码。 30多个tRNA基因都由叶绿体DNA编码
且以细胞质遗传
的方式稳定遗传
2. 叶绿体的遗传特点
叶绿体具有遗传自主性: 既能自我复制,又能独立表达(决定性状表现)
叶绿体遗传的自主性是相对的:
在某些情况下受核基因影响(甚至产生细胞质 基因突变)
*叶绿体遗传的分子基础
1962年,由Kis和Plant首先明确肯定植物细胞的 叶绿体中含有DNA。
N:具有正常的细胞质 (可育胞质)
S:具有不育的细胞质 (不育胞质)
RR:显性纯合可育核基因型 Rr:杂合可育核基因型 rr:隐性纯合不育核基因型
细胞核的遗传类型:可育(R) 对不育(r)为显性
不育系
以不育系作为母本,5种可育类型作为父本,后代
的育性遗传表现如图:
1、
不育系——S(rr) 恢复系——N(RR)4
核外遗传
第一节 核外遗传的概念和特点
核外遗传:
由位于核或类核体以外的遗传物质所
决定的遗传现象和遗传规律,也称为细胞 质遗传,非孟德尔遗传.
细胞质遗传的实例和特点
紫茉莉的花斑遗传 1909年由Carl Corrans 发现(irabilis jalapa)
紫茉莉花斑植物杂交的结果 母本枝条表型 白色 父本枝条表型 白色 绿色 花斑 绿色 白色 绿色 花斑 花斑 白色 绿色 花斑 后代表型 白色 白色 白色 绿色 绿色 绿色 花斑,绿色,白色 花斑,绿色,白色 花斑,绿色,白色
人类线粒体疾病
典型的母系遗传,
只有女性才能传递
线粒体疾病
同一家系中不同患
者表现症状有差 异—(异质性)
2. 线粒体遗传的特点
与质体的遗传相比,线粒体基因组的遗传与核基 因组间的关系更紧密
一方面,线粒体在遗传具有半自主性,能够自
我复制、转录和翻译表达 (决定性状表现)
另一方面,线粒体基因组编码线粒体中不到
这种由母体基因型决定子代性状表现的母性影响又叫前定
作用(per-determination)、或延迟遗传(delayed inheritance)。
螺类卵裂的方式示意图
1、2为第一次卵裂,