遗传学第八章核外遗传分析
第八章-核外遗传

第八章核外遗传重点:掌握核基因和细胞质基因的遗传特点➢细胞质基因➢母性影响➢核外遗传的性质与特点➢细胞内敏感性物质的遗传➢线粒体的遗传方式及其分子基础第一节细胞质基因染色体以外的遗传因子所决定的遗传现象,在真核生物中称为核外遗传或细胞质遗传(extranuclear inheritance或cytoplasmic inheritnce)。
细胞质中的线粒体、叶绿体、共生体、游离体〔中心球〕、动质体〔某些鞭毛原生动物的线粒体很特殊,它与一个DNA的特异结构联系在一起,称之为动质体〕均含有DNA。
细胞质基因和核基因的异同点:相同点:1、二者都是以半保留方式进行自我复制。
2、蛋白质合成中,二者都是首先以mRNA形式进行转录,而后在核糖体上进行翻译的,并且线粒体本身具有独立的核糖体。
3、二者的基因都可能发生突变,而且诱变的因素也是相同的,并且二者产生的基因突变都是可能遗传的。
不同点:1、细胞质基因的遗传不遵守孟得尔定律。
2、细胞质基因人工诱变突变频率比核基因诱变突变的频率明显的高,并且有专一性。
3、胞质基因的遗传传递不通过雄性配子,而是由卵子传递的。
4、细胞质基因只有个别的游离基因能与核基因,在细胞分裂中同步分裂,而绝大多数细胞质基因是呈无规律的别离和重组的。
5、有些细胞质基因不但具有遗传属性,而且还有接触感染的属性。
第二节母性影响一、短暂的母性影响在正反交情况下,子代某些性状相似于其雌性亲本的现象,有的是由于细胞质遗传因子传递的结果,属于核外遗传的范畴。
但有的却是由于母体中核基因的某些产物积累在卵细胞的细胞质中,使子代表型不由自身的基因所决定而与母本表型相同的遗传现象,则称为母性影响(maternal inheritance)。
母性影响有两种:一种是短暂的,只影响子代个体的幼龄期;另一种是持久的,影响子代个体终生。
欧洲麦蛾(Ephestia kuhuniella)野生型幼虫的皮中含有色素,成虫复眼为深褐色由于基因A使幼虫着色,卵母细胞中有A基因存在,因而经减数分裂产生的卵,不管基因型是A或a它们的细胞质中都含有A基因产物,即色素的前体以及由前体所合成的色素。
遗传学经典核外遗传
线粒体基因组
mtDNA的基因组成 (1)闭合环状DNA (2) 基因数目和排列顺序相同
(3)有D环和2个复制起始点 (4)基因间没有间隔,因此每个基因不可
核密码 终止 Arg Ile Ile Arg 终止 终止 Ile Leu Leu 终止 终止
核糖体的差异
表 20-4 人类和酵母线粒体核糖体和细胞质核糖体的大小
成分
人类(HeLa)
mt
cyto
酵母
mt
Cyto
核糖体
60S
74S
75S
80S
大亚基
45S
60S
53S
60S
小亚基
35S
40S
35S
40S
能都有自己的起动子 (5)某些蛋白质的密码子与核基因通用
密码子不同; (6)mtDNA主要编码rRNA和tRNA分子,氧化
呼吸所需要的酶类少部分亚基。
酵母的小菌落突变
酵母菌可以进行有性生殖,与链孢霉类似。 正常菌落能自发形成部分小菌落(1%2%),化学诱变剂能诱导小菌落。 小菌落:细胞生长缓慢,长成很小的菌落。 缺少细胞色素a、b以及细胞色素c氧化酶。 稳定遗传。
线粒体中密码子的改变
生物 共同(人、果蝇、酵母) 哺乳动物 哺乳动物 果蝇 果蝇 纤毛虫 另一类纤毛虫 酵母 酵母 酵母 酵母 支原体
密码子 UGA AGA/AGG AUA AUA AGA UAA/UAG UGA/UAG AUA CCA CUG UAG UGA
遗传学第八章 核外遗传分析课件

2、 性比(sex-ratio,SR)因子
◇SR因子是胞质中的一种原生动物,在雌蝇 和雄蝇中都能发现,但对发育中的雄性幼 虫是致死的,所以后代中雌蝇比例远大于 雄蝇比例。
◇将SR雌蝇的卵细胞质注入正常雌蝇可诱导 SR现象。
◇有证据表明,产生雄性致死毒素的可能是原 生动物内的病毒。
基因型与表型的关系
细胞质基因 正常(N) 不育(S)
核基因型
RfRf(可育) Rfrf(可育) rfrf(不育) N(RfRf)可育 N(Rfrf)可育 N(rfrf)可育 S(RfRf)可育 S(Rfrf)可育 S(rfrf)不育
遗传学 第八章 核外遗传分析
(二)可能的遗传机制 1、线粒体与雄性不育的关系 2、叶绿体与雄性不育的关系
遗传学 第八章 核外遗传分析
遗传学 第八章 核外遗传分析
• KSS综合征(Keams-Sayre Syndrome)是多 系统线粒体病,主要症状为眼肌麻痹和色素性视 网膜炎。患者骨骼肌细胞mtDNA有2.0kb7.0kb的缺失。 遗传学 第八章 核外遗传分析
五、叶绿体遗传及其分子基础
(一)衣藻的叶绿体遗传
◇ 衣藻细胞中只有一个叶绿体,约含50个 拷贝的环状ds-DNA分子。 ◇ 不同交配型(mt+,mt-)的单倍体衣藻 杂交,形成短暂2n时期,进行减数分裂。 ◇ 虽然杂交双方融合时为合子提供等量细 胞质,但叶绿体只由mt+方传递,表现单亲 遗传。
遗传学 第八章 核外遗传分析
遗传学 第八章 核外遗传分析
(二)叶绿体遗传的分子基础 1、叶绿体基因组
大小:环状双链DNA分子。大小120-190kb。 其基因序列中不含5-甲基胞嘧啶。
◆ cpDNA编码约100种蛋白质和RNAs,包 括45个编码RNA的基因,27个编码与基因表达 有关的蛋白的基因,18个编码类囊体膜的蛋白基 因和10个与电子传递功能有关的基因。
《核外遗传分析》课件

核外遗传的特点
核外基因组结构简单,基 因密度高,复制和转录过 程相对简单。
核外基因组具有母系遗传 的特点,即线粒体基因组 和叶绿体基因组只来自母 本。
核外基因组在进化上相对 保守,但在某些物种中也 可能发生基因重组和突变 。
核外遗传分析的意义
核外遗传分析有助于深入了解生 物的进化历程和物种起源。
详细描述
表观遗传学分析在核外遗传分析中扮演着重要的角色。表观遗传学标记如DNA甲基化和 组蛋白修饰等可以影响基因的表达水平,进而影响生物体的表型。通过分析这些表观遗 传学标记,可以深入了解基因表达的调控机制,为疾病诊断和治疗提供新的思路和方法
。
转录组学分析
总结词
转录组学分析主要研究基因转录本的种类和丰度,通过分析转录本的变化,揭示基因表达的动态变化 和调控机制。
和思路。
02
个体化用药
根据个体的核外遗传变异情况,可以制定针对个体的个性化用药方案,
提高药物的疗效并降低副作用。
03
药物反应差异研究
不同个体对同一种药物的反应可能存在差异,这种差异可能与个体的核
外遗传变异有关,通过研究这种关系可以更好地理解药物反应的个体差
异。
个体化医疗的核外遗传分析应用
精准医疗
基于个体的核外遗传变异情况,可以为患者提供精准、个性化的 治疗方案,提高治疗效果。
技术发展的挑战
技术更新换代
01
核外遗传分析技术不断发展,需要不断更新知识和技术,以适
应新的分析需求。
技术局限性
02
目前核外遗传分析技术仍存在一定的局限性,如检测灵敏度、
特异性等方面仍有待提高。
技术普及
03
如何将核外遗传分析技术普及到临床实践中,提高其在医学研
分子遗传学核外遗传

染色体外DNA可以在细胞内自 主复制和传播,影响基因的表达
和细胞功能。
染色体外DNA的变异和扩增可 以导致基因组不稳定和疾病的发
生。
03
核外遗传物质的特性
遗传方式的多样性
01
核外遗传物质包括线粒体DNA和叶绿体DNA,它们分别通过母 系和父系遗传。
02
线粒体DNA和叶绿体DNA的基因组结构简单,基因数目较少,
生物燃料
利用核外遗传物质改良微 生物,提高微生物产乙醇 等生物燃料的能力,降低 生产成本和提高产量。
05
核外遗传物质的挑战与前 景
核外遗传物质研究的挑战
技术难度
核外遗传物质的研究需要高精度 的技术手段,如测序技术、基因 编辑等,这些技术目前仍处于不 断发展和完善阶段,存在一定的
技术难度。
样本获取
分子遗传学的重要性
基础研究
分子遗传学为生命科学领域的基 础研究提供了重要的理论支撑, 推动了生物学、医学、农学等领
域的发展。
医学应用
分子遗传学在医学领域的应用广泛, 如疾病诊断、治疗和预防等方面, 为人类健康提供了有力支持。
农业实践
分子遗传学在农业领域的应用,如 品种改良、抗逆性育种等方面,提 高了农业生产效率和可持续性。
核外遗传物质的研究需要大量的 样本,包括细胞、组织等,这些 样本的获取需要耗费大量时间和 精力,且存在一定的伦理问题。
数据解读
核外遗传物质的研究会产生大量 的数据,如何解读这些数据,挖 掘其中的生物学意义,需要具备 深厚的生物学和数据分析知识。
核外遗传物质研究的前景
疾病诊断与治疗
随着核外遗传物质研究的深入,人们将更加了解疾病的发病机制, 为疾病的诊断和治疗提供新的思路和方法。
遗传学名词解释(答案)

名词解释第一章绪论遗传学:是研究生物遗传和变异的科学,是生物学中一门十分重要的理论科学,直接探索生命起源和进化的机理。
同时它又是一门紧密联系生产实际的基础科学,是指导植物、动物和微生物育种工作的理论基础;并与医学和人民保健等方面有着密切的关系。
遗传:是指亲代与子代相似的现象。
如种瓜得瓜、种豆得豆。
变异:是指亲代与子代之间、子代个体之间存在着不同程度差异的现象。
如高秆植物品种可能产生矮杆植株,一卵双生的兄弟也不可能完全一样。
第二章遗传的细胞学基础染色质:是指染色体在细胞分裂的间期所表现的形态,呈纤细的丝状结构,含有许多基因的自主复制核酸分子。
染色体:在细胞分裂时期,在细胞核中容易被碱性染料染色、具有一定数目和形态结构的的杆状体。
(染色体:指任何一种基因或遗传信息的特定线性序列的连锁结构。
)染色单体:由染色体复制后并彼此靠在一起,由一个着丝点连接在一起的姐妹染色单体。
姐妹染色单体:二价体中的同一各染色体的两个染色单体,互称姐妹染色单体,它们是间期同一染色体复制所得。
非姐妹染色单体:单体二价体的不同染色体之间的染色单体互称非姐妹染色单体,它们是同源染色体这些间期各自复制所得。
联会:减数分裂中,同源染色体的配对过程。
同源染色体:大小,形态和结构相同,功能相似的一对染色体。
非同源染色体:形态和结构不同的各对染色体互称为非同源染色体。
有丝分裂:包含两个紧密相连的过程:核分裂和质分裂。
即细胞分裂为二,各含有一个核。
分裂过程包括四个时期:前期、中期、后期、末期。
在分裂过程中经过染色体有规律的和准确的分裂,而且在分裂中有纺锤丝的出现,故称有丝分裂。
减数分裂:又称成熟分裂,是在性母细胞成熟时,配子形成过程中所发生的一种特殊的有丝分裂。
它使体细胞染色体数目减半。
它含两次分裂,第一次是减数的,第二次是等数的。
双受精:授粉后,一个精核(n)与卵细胞(n)受精结合为合子(2n),将来发育成胚。
同时另一精核(n)与两个极核(n+n)受精结合为胚乳核(3n),将来发育成胚乳。
第八章 核外遗传

外包双层膜、直径为0.2~0.4μm的小颗粒,含DNA、
RNA和酶。
卡巴粒的rRNA能与E. coli DNA杂交,但不能与
草履虫的DNA杂交。
认为卡巴粒是一种共生型的具有螺旋结构的放毒
型细菌。
其它放毒性颗粒:γ粒、λ粒、μ粒。
二、果蝇的感染性遗传
果蝇对CO2的敏感性的遗传
果蝇“性比(SR)失常”
突变型a
不产生犬尿氨酸,幼虫皮肤无色,成 虫复眼红色。
(能合成犬尿氨酸)
麦粉蛾的色素遗传中的母体影响
无色 结果同一般测交
是短暂的,只影响子代的幼龄期。
2、持久的母体影响
(d)
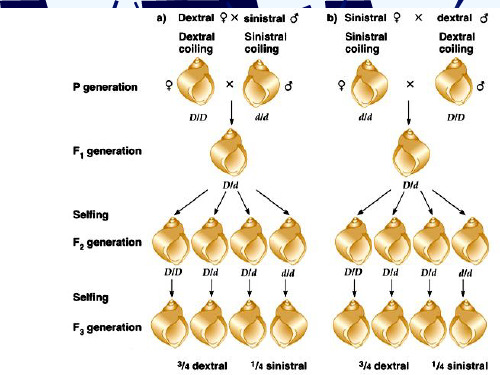
椎实螺的外壳
(D)
椎实螺外壳旋向的遗传
母体影响
母体 影响 母体中核基因D的产物积累在卵母细胞的 细胞质中,使F2中的dd个体也表现右旋.
线粒体,小菌落消失。
二、线粒体基因组
线粒体基因组的一般性质
线粒体DNA的组成和蛋白
质合成
人类的线粒体基因病
(一) 线粒体基因组的一般性质
1962年,由KIS等发现线 粒 体 ( mitochondrion ) 内有DNA(mtDNA), 与核内DNA明显不同
环状(线状)、裸露
不同生物线粒体DNA的比较
酵母菌
固体培养基上通气培养
大菌落
缺少细胞色 素a,b和细 小菌落 胞色素c氧 1-2% 化酶—呼吸 缺陷突变株
大菌落
小菌落 1-2%
小菌落
都为正常型菌落
解 释
小菌落的形成:线粒体DNA的遗传变
异。
小菌落与正常的酵母杂交:细胞质融
核外遗传讲解

MTDNA的特点
在组成上,绝大多数mtDNA中没有重复核苷 酸序列(这是mtDNA一级结构的重要特点)。 mtDNA的浮力密度较低。 mtDNA的碱基成分中G、C含量比A、T少。 mtDNA的两条单链密度不同,一条称为重链 (H链),另一条称为轻链(L链)。
mtDNA单个拷贝非常小,仅有核DNA的十万分 之一。
以S(RR)作母本与五种可育个体杂交结果
♀ ♂ F1 S(rr)不育×S(RR)可育 S(Rr)可育 S(rr)不育×S(Rr)可育 S(rr)不育 S(rr)不育×N(Rr)可育 S(Rr)可育 S(rr)不育×N(RR)可育 S(Rr)可育 S(rr)不育×N(rr)可育 S(rr)不育 上术各种杂交组合可归纳为三种情况 保持系 ——N(rr)具有保持不育性在世代中稳定遗传的 能力,称为保持系 。 不育系——S(rr)能被N(rr)所保持,使其后代出现全 部稳定不育的个体,称为不育系。 恢复系——N(RR)或S(RR)具有恢复育性的能力,称为 恢复系。
2.
叶绿体DNA的物理图谱及基因定位
(1) 4.5S、5S、16S、23SrDNA(分布在反向重复序列区);
(2) 30多个tRNA编码序列——4个在反向重复区,其中2个 在16S和23S rDNA之间、其余分散分布; (3)mRNA: 核糖体蛋白(1/3,20)、Rubisco大亚基、PSI 和PSII(部分)、ATPase、RNA多聚酶(部分)、电子传递蛋 白等。
二、叶绿体遗传的分子基础
叶绿体基因组特点 1.经DNA序列分析含有两个反向重复序列(IRS)。 2.反向重复序列在复性时形成互补双链,它们之间非重复 序列形成两个大小不等的环。 3.有些cpDNA有内含子,cpDNA无5„-甲基胞嘧啶。 4.叶绿体DNA能自我复制。 5.叶绿体基因组含有自己的转录翻译系统。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
可编辑ppt
1
紫茉莉(Mirabilis jalapa)植株的花斑
(variegation)的遗传
可编辑ppt
2
二、细胞内敏感性物质的遗传
• 感染性遗传是一种通过细胞质传递的遗传方式, 由入侵真核生物细胞质的微生物或生物粒子引起。 一般通过母方细胞质传递到后代,赋予特殊的表型。
(一)草履虫的放毒型遗传
可编辑ppt
3
草履虫的有性生殖方式
• ◇接合(conjugation):经过接合,由 纯合体变成杂合体。
• ◇自体受精(autogamy):由杂合体变 成纯合体。
可编辑ppt
4
可编辑ppt
5
可编辑ppt
6
(二)果蝇的感染性遗传
1、 对CO2敏感的б因子 ◇б因子是小于Kappa颗粒的病毒,存在于果
核基因型
RfRf(可育) Rfrf(可育) rfrf(不育) N(RfRf)可育 N(Rfrf)可育 N(rfrf)可育 S(RfRf)可育 S(Rfrf)可育 S(rfrf)不育
症状:为突发的双侧性中央视力丧失,相关的四
种突变均扰乱正常的氧化磷酸化。50%以上的病 例是由于线粒体上编码NADH脱氢酶的基因的 No.11 778位G→A引起。许多LHON病例无家 族史。
• KSS综合征(Keams-Sayre Syndrome)是多
系统线粒体病,主要症状为眼肌麻痹和色素性视
网膜炎。患者骨骼肌细胞mtDNA有2.0kb-
● 雄性不育(male sterility):雄蕊 发育不正常,不能产生有功能的花粉,但 雌蕊发育正常,能接受正常花粉而受精结 实。
● 应用价值:在杂种优势利用中,省去 大量去雄工作,保证杂交种子的纯度。
可编辑ppt
28
(一)植物雄性不育的类型 1、核不育 2、核-质互作不育型
基因型与表型的关系
细胞质基因 正常(N) 不育(S)
细胞学证据:异常线粒体内的碎红纤维和仲 晶(para crystalliance)排列。
分子机制:主要由mtDNA tRNA lys基因第8344
位的A→G引起。
可编辑ppt
18
可编辑ppt
19
• leber氏遗传性视神经病(leberˊs hereditary optic neuropathy,LHON)也表现母系遗传 和线粒体缺损。
白,9种ATP合成酶,16种核糖体合成蛋白,29
种未知功能蛋白等。
可编辑ppt
14
可编辑ppt
15
线粒体基因遗传密码的变化:
可编辑ppt
16
2、mtDNA的结构特点 3、人类线粒体基因病
人类mtDNA编码的基因产物:13种氧化磷 酸化所需蛋白质;22种翻译所需tRNAs和2种翻 译所需rRNAs。mtDNA异常会引起多种线粒体 病。
蝇的胞质中。
◇胞质中含有б因子的果蝇经CO2麻痹后,不能 象抗性果蝇一样当放回正常空气中时恢复正常, 而是长期麻痹并死亡。
◇敏感雌蝇可将б因子和CO2敏感的性状传递到 后代。
◇将敏感果蝇的提取物注入抗性果蝇能将其诱导 为敏感果蝇。
可编辑ppt
7
2、 性比(sex-ratio,SR)因子
◇SR因子是胞质中的一种原生动物,在雌蝇 和雄蝇中都能发现,但对发育中的雄性幼 虫是致死的,所以后代中雌蝇比例远大于 雄蝇比例。
第八章 核外遗传分析
一、核外遗传的性质与特点 ◆ 性状由细胞器基因决定,基因不在染色体
上,遗传方式是非孟德尔式的。 ◆ 细胞器位于胞质中,杂交时主要由母方提
供遗细 传胞 (质ma,teFr1通na常l i只nh表er现it母an方ce的)性状,即母系 ◆ 细胞器基因在细胞分裂中不遵从染色体上
的分离原则,杂交后代一般不出现孟德尔分离 比。
7.0kb的缺失。
可编辑ppt
20
五、叶绿体遗传及其分子基础
(一)衣藻的叶绿体遗传
◇ 衣藻细胞中只有一个叶绿体,约含50个 拷贝的环状ds-DNA分子。
◇ 不同交配型(mt+,mt-)的单倍体衣藻 杂交,形成短暂2n时期,进行减数分裂。
◇ 虽然杂交双方融合时为合子提供等量细 胞质,但叶绿体只由mt+方传递,表现单 亲遗传。
◇ mtDNA突变与相关疾病
可编辑ppt
17
• 肌痉挛性癫痫与破损性红肌纤维病 (myoclonic epilepsy and ragged red fiber disease,MERRF)是一种母系遗传 的线粒体脑病,10-20岁发病。
症状:主要为骨骼肌的不自主痉挛,全身性 的强直痉挛发作和失神,晚期出现精神异 常。
◇将SR雌蝇的卵细胞质注入正常雌蝇可诱导 SR现象。
◇有证据表明,产生雄性致死毒素的可能是 原生动物内的病毒。
可编辑ppt
8
三、母性影响
(一)短暂的母体影响(欧洲麦蛾)
可编辑ppt
9
(二)持久的母性影响(椎实螺)
可编辑ppt
10
四、线粒体遗传及其分子基础 (一)酵母小菌落突变
可编辑ppt
11
可编辑ppt
可编辑ppt
23
可编辑ppt
24
结构特点:
地钱M.polymorpha cp基因组为
121kb,是第一个测定全序的cpDNA,含 有92个蛋白编码基因,36个rRNA和tRNA 编码基因
可编辑ppt
25
可编辑ppt
26
• 2、叶绿体遗传系统与核遗传系统的关系
可编辑ppt
27
六、植物雄性不育
◇ 酿酒酵母mt基因组为75kb的环状dsDNA, 有结构基因,rRNA基因和tRNA基因,其中编码 细胞色素C和细胞色素氧化酶的基因是断裂的, 长的内含钱M.polymorpha mt基因组为186kb,
是最小的植物mt基因组之一,但编码的基因数远
比非植物mt基因组多,包括12种电子传递链蛋
可编辑ppt
21
可编辑ppt
22
(二)叶绿体遗传的分子基础
1、叶绿体基因组
大小:环状双链DNA分子。大小120-190kb。
其基因序列中不含5-甲基胞嘧啶。
◆ cpDNA编码约100种蛋白质和RNAs,包 括45个编码RNA的基因,27个编码与基因表达 有关的蛋白的基因,18个编码类囊体膜的蛋白基 因和10个与电子传递功能有关的基因。
12
(二)线粒体基因组
1、线粒体基因组的一般性质
高等植物:200-2500kb
酵母: 75kb
脉孢菌: 60kb
牛:
17533bp
人类: 16569bp
可编辑ppt
13
三种线粒体基因组的特点:
◇ 人mt基因组为16.5kb的环状dsDNA,有22 个tRNA基因,2个rRNA基因和13个蛋白质编码 区。