大肠杆菌高效表达重组蛋白策略
工程菌

两阶段培养法
为了提高质粒稳定性,工程菌培养采 用两阶段培养法: ⑴先使菌体生长至一定密度 ⑵再诱导外源基因的表达
⑴使菌体生长至一定密度 在生长阶段使外源基因处于阻遏状态,避 免由于基因表达造成基因不稳定性遗传, 使质粒稳定地遗传。 ⑵诱导外源基因的表达 在获得需要的菌体后,再去阻遏或诱导外源 基因的表达。
5.pH
• pH对细胞的正常生长和外源蛋白的高效表 达都有影响,所以应根据工程菌的生长和 代谢情况,对pH进行适当的调节。
• 如重组Lactococcus lactis ctis(pIL252)对生长和质粒稳定性 的最适pH值分别为6.39和6.41。
选择合适的宿主菌
一般情况下,质粒的结构不稳定性可 以通过宿主菌株的谨慎和恰当的选用来消 除。 宿主菌通常有苏云金芽孢杆菌、根瘤 菌、酒精酵母和大肠杆菌等。 大肠杆菌由于具有生长周期短、方法 简便、价格低廉、表达水平高诸多优点成 为基因工程的首选表达系统。
• 在大肠杆菌表达系统中通常可以选用T7、 PRP、Ptac等强启动子,通过对翻译增强 序列和转录终止序列的改进和优化、实现 外源基因的高效表达。 • 可以按照大肠杆菌系统偏爱的密码子和 mRNA二级结构进行目的基因的设计和合成, 获得外源基因的高效表达。 • 在大肠杆菌表达系统中采用F质粒的ccd毒 素-抗毒素联合系统可在细胞分裂时杀死丢 失质粒的细胞,从而维持高的质粒稳定性。
1. 外源基因的拷贝数
• 一般来说细菌内基因拷贝数增加,基因 的表达产物也增加。外源基因是克隆到 载体上的,因此载体在宿主细胞中的拷 贝数就直接关系到外源基因的拷贝数。 将外源基因克隆到高拷贝数的表达质粒 上,含外源基因的重组表达质粒拷贝数 的增加,必然导致外源基因拷贝数的增 高,这对于外源基因的总体表达水平非 常有用。
大肠杆菌表达体系的特点

大肠杆菌表达体系的特点
大肠杆菌表达体系是一种常用的重组蛋白表达方法,具有以下特点:
1. 简单易用:大肠杆菌是一种常见的细菌,易于培养和操作。
其表达体系基于质粒介导的转化和表达,具有操作简单、成本低廉的优点。
2. 高表达水平:大肠杆菌表达体系能够实现高表达水平,通常可以达到10-50%的总蛋白含量。
这一特点使其成为生物制药和科学研究领域中最受欢迎的表达体系之一。
3. 多种表达宿主:大肠杆菌表达体系有多种表达宿主,包括
BL21(DE3)、Rosetta(DE3)、Origami(DE3)等。
这些表达宿主具有不同的特点,能够适应不同的表达需求。
4. 可定制化:大肠杆菌表达体系可以通过基因工程技术进行改造,实现蛋白质的定制化表达。
例如,可以通过融合表达标签、选择性培养、调控表达等方式来优化表达效果。
5. 可用于生物制药:大肠杆菌表达体系可以用于制备多种蛋白药物,如重组人胰岛素、干扰素、白介素等。
这些蛋白药物已经被广泛应用于临床治疗和研究领域。
总之,大肠杆菌表达体系是一种快速、高效、可定制化的表达系统,已经成为蛋白质表达和生物制药领域中最常用的表达系统之一。
- 1 -。
大肠杆菌中高效表达外源蛋白的策略
在大肠杆菌中高效表达外源蛋白的策略赵军侯云德(病毒基因工程国家重点实验室 100052 北京)本世纪60至70年代对大肠杆菌的研究使之成为自然界中最普遍为人们所认识的生物体。
大肠杆菌具有两个显着特征:操作简单和能在廉价的培养基中高密度培养,它的这些特征加上十多年外源基因表达的经验使其在大多数科研应用中成为高效表达异源蛋白最常用的原核表达系统。
尽管大肠杆菌有众多的优点,但并非每一种基因都能在其中有效表达。
这归因于每种基因都有其独特的结构、mRNA的稳定性和翻译效率、蛋白质折叠的难易程度、宿主细胞蛋白酶对蛋白质的降解、外源基因和E.coli在密码子利用上的主要差别以及蛋白质对宿主的潜在毒性等等。
但知识的大量积累还是有助于为表达方面某些特定的困难提供一般的解决方法。
大肠杆菌作为表达系统的主要障碍包括:不能象真核蛋白那样进行翻译后修饰、缺乏将蛋白质有效释放到培养基中的分泌机制和充分形成二硫键的能力。
另一方面,许多真核蛋白在非糖基化的形式下能保留其生物学活性,因而也就可以用大肠杆菌来表达。
如何实现外源基因在原核细胞中的有效表达,自60年代以来,对影响外源基因在其表达体系中表达效率的各个因素作了大量实验研究,并有多篇归纳性综述发表[1,2,3]。
国内针对外源基因在原核细胞中高效表达的关键因素,构建了高效表达载体[4],并在此基础上成功表达了一系列细胞因子的基因[5,6,7]。
我们在分析了国内外有关在原核系统中表达蛋白的实验资料的基础上,对在大肠杆菌中高效表达外源蛋白的策略所涉及的内容进行全面的总结,以期有助于我国在这方面的研究。
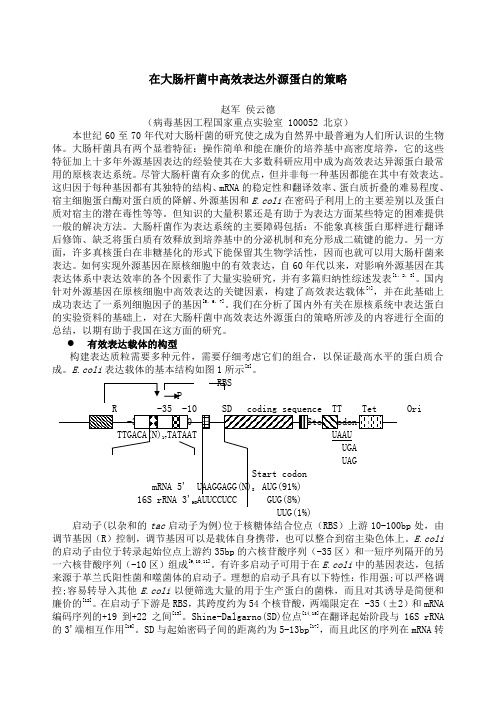
有效表达载体的构型构建表达质粒需要多种元件,需要仔细考虑它们的组合,以保证最高水平的蛋白质合成。
E.coli表达载体的基本结构如图1所示[8]。
RBSP启动子(以杂和的tac启动子为例)位于核糖体结合位点(RBS)上游10-100bp处,由调节基因(R)控制,调节基因可以是载体自身携带,也可以整合到宿主染色体上。
在大肠杆菌中表达重组蛋白的流程

在大肠杆菌中表达重组蛋白的流程
在大肠杆菌中表达重组蛋白的流程通常包括以下步骤:
1. 克隆:首先需要将目标基因克隆到适当的表达载体中。
这可以通过PCR扩增目标基因,然后将其与表达载体连接,形成重组质粒。
2. 转化:将重组质粒转化到大肠杆菌细胞中。
可以使用化学方法(如热冲击法)或电穿孔法将质粒导入细胞。
3. 选择:转化后,将细胞分散在含有适当抗生素的琼脂平板上培养。
只有带有重组质粒的细胞能够存活并形成菌落。
4. 培养:将含有重组细胞的培养液转移到适当的培养基中,并在适当的条件下培养。
这可能包括调节温度、pH值和搅拌速度等。
5. 表达:在培养期间,目标基因会被大肠杆菌细胞转录和翻译为蛋白质。
使用适当的启动子和调控序列,可实现目标蛋白的高效表达。
6. 细胞破碎:一旦细胞达到最佳表达水平,就需要破碎细胞以释放目标蛋白。
这可以通过多种方法实现,如超声波、高压破碎或化学方法。
7. 纯化:通过使用各种分离和纯化技术(如亲和层析、凝胶过滤、离子交换层析等),从细胞裂解液中纯化目标蛋白。
以上是在大肠杆菌中表达重组蛋白的一般流程。
具体的步骤和条件可能因实验设计和目标蛋白的特性而有所不同。
大肠杆菌重组蛋白表达流程

大肠杆菌重组蛋白表达流程大肠杆菌重组蛋白表达流程主要包括以下几个步骤:1. 选择合适的表达载体:通常选择含有启动子、转录终止子、选择标记和适当的表达调控元件的表达载体。
启动子用于驱动基因转录,转录终止子用于确定转录产物的结束位置,选择标记有助于筛选含有目的基因的转化子,而表达调控元件可以调节基因的表达水平。
2. 构建表达载体:将目的基因插入表达载体中,构建成重组表达载体。
在此过程中,需要考虑目的基因的orientation(方向)、阅读框(ORF)以及表达调控元件的活性等因素。
3. 转化大肠杆菌:将构建好的重组表达载体转化到大肠杆菌中。
转化方法有多种,如化学法(如CaCl2法)、电转化、热激转化等。
转化后,大肠杆菌吸收了外源DNA,成为重组菌株。
4. 筛选重组菌株:在含有选择性抗生素的培养基上培养转化后的菌落,筛选出含有目的基因的重组菌株。
此外,可以通过鉴定菌落的形态、颜色等特征进行初步筛选。
5. 诱导表达:将筛选出的重组菌株接种到含有诱导剂(如IPTG)的培养基中,诱导目的基因的表达。
诱导剂IPTG可以与表达载体中的启动子结合,增强基因转录和翻译的效率。
6. 收集和纯化重组蛋白:诱导表达后,菌体中会含有目的蛋白。
可以通过离心、破碎细胞、柱层析等方法分离和纯化重组蛋白。
常用的纯化标签有His标签、GST标签等,这些标签可以帮助分离和纯化目的蛋白。
7. 蛋白活性检测和应用:对纯化的重组蛋白进行活性检测,如酶活测定、蛋白互作实验等。
确认蛋白活性后,可应用于生物学研究、药物研发等领域。
需要注意的是,大肠杆菌重组蛋白表达过程中可能会遇到表达量低、蛋白包涵体等问题。
为了解决这些问题,可以尝试优化表达载体、改变诱导条件、使用融合标签等策略。
重组人胶原蛋白肽在大肠杆菌中的高效表达

重组人胶原蛋白肽在大肠杆菌中的高效表达陈光;吴铭;王刚;孙旸【摘要】目的:构建含有胶原蛋白基因的原核表达载体pET32a-CP6,并转入大肠杆菌BL21 (DE3)中进行高效表达,为获得大量可溶的胶原蛋白肽提供可靠依据.方法:将重组表达载体pET32a-CP6转化到大肠杆菌BL21中,以IPTG为诱导剂,对温度、接种量、诱导时机、IPTG诱导浓度等各种发酵参数进行优化,筛选高效表达的发酵条件,并通过Ni-NTA亲和层析进行纯化分析.结果:菌株在37℃、接种量为2%、摇瓶培养约2.5h后,加入终浓度为0.5mmol· L-1的IPTG,37℃诱导5h,收获的蛋白产量最高为31.52 mg· L-1,Western blotting分析该表达产物与人COL6A2单克隆抗体有特异结合能力.结论:优化工程菌高效表达重组人胶原蛋白,纯化重组蛋白.【期刊名称】《吉林大学学报(医学版)》【年(卷),期】2011(037)004【总页数】5页(P656-660)【关键词】重组胶原蛋白肽;重组蛋白纯化;大肠杆菌【作者】陈光;吴铭;王刚;孙旸【作者单位】吉林农业大学生命科学院生物物理实验室,吉林长春130118;吉林农业大学生命科学院生物物理实验室,吉林长春130118;吉林农业大学生命科学院生物物理实验室,吉林长春130118;吉林农业大学生命科学院生物物理实验室,吉林长春130118【正文语种】中文【中图分类】Q78胶原蛋白是由动物的成纤维细胞合成的一种生物高分子,广泛存在于动物骨、韧带、软骨、皮肤及其他结缔组织中,是细胞外基质的主要成分[1]。
由于其特有的生理活性和丰富的营养价值,被广泛地应用于医学、美容、化妆品、食品等领域。
目前胶原蛋白的制备大多采用常规方法,通常以含有结缔组织的屠宰陆生动物废弃物为原料,对其进行酸、碱和酶处理来提取胶原蛋白,但该方法提取的蛋白具有非水溶性、病毒传染性、异体免疫排斥性等缺点,限制了胶原蛋白许多潜在用途的开发[2]。
基因在大肠杆菌、酵母中的高效的表达

c. 启动子与克隆基因间的距离对基因表达的影响
研究表明启动子和目的基因间的距离对基因的 表达效率影响很大,所以在构建新的表达载体时要考 虑到这一因素的影响。另外,在克隆基因的末端要就 近插入有效的终止子序列,否则会导致细胞能量的大 量消耗,或是形成不应有的二级结构,最终影响的目 的基因的表达效率。
影响目的基因在甲醇酵母中表达的因素
1.目的基因的特性 2.表达框的染色体整合位点与基因拷贝数 3.宿主的甲醇利用表型 4.分泌信号 5.产物稳定性 6.翻译后修饰
பைடு நூலகம்
b. 翻译起始序列对表达效率的影响
mRNA的有效翻译依赖于核糖体和其的稳定结 合,大肠杆菌的mRNA序列中,核糖体的结合位点是 起始密码子AUG和其上游的SD序列。所谓SD序列就 是由Shine-Dalgarno首先提出的一种位于位于起始密 码子上游的一段保守序列,为细菌核糖体有效结合和 翻译起始所必需。一般SD序列的长度约为3-9bp,位 于起始密码子上游3-11碱基的位置,它与16S核糖体 RNA的3‘端互补,控制了翻译的起始。 5’--AGGAGGUXXXAUG--- mRNA 3’AUUCCUCCACUAG----- 16S rRNA3’ 末端
构建表达载体的策略
⑴将真核基因克隆到一个强大的原核启动子和SD序列
的下游,使得真核基因处于原核调控体系中。 ⑵采用真核基因的cDNA序列作为构建表达载体的目的 基因,这样就解决了原核细胞没有RNA剪接功能的 问题。
⑶构建载体时,将真核基因插在几个原核密码子的后 面,翻译后就得到了原核多肽和真核多肽的融合 蛋白,这样就可以避免被原核蛋白酶的识别和降 解,最后可以将融合多肽切除。
3. mRNA合成后穿过核膜进入细胞质中后才进行翻译 工作,而且通常都有复杂的成熟和剪接过程; 4. 基因的启动子区和原核基因差异很大,而且有增强 子序列存在。
大肠杆菌表达

大肠杆菌表达引言大肠杆菌(Escherichia coli)是一种常见的细菌,广泛应用于基因工程和蛋白质表达领域。
大肠杆菌表达系统具有高效、经济且易于操作的特点,因此被广泛用于重组蛋白的生产。
本文将介绍大肠杆菌表达系统的基本原理及其在蛋白质表达中的应用。
大肠杆菌表达系统的基本原理大肠杆菌表达系统采用重组DNA技术,将外源基因插入到大肠杆菌的表达载体中。
表达载体通常包含一个启动子、一个转录终止子、一个选择性抗生素抗性基因和一个参考基因。
启动子能够促使外源基因的转录,转录终止子能够终止转录过程,选择性抗生素抗性基因则能够确保只有带有外源基因的细菌存活下来。
参考基因用于对比表达水平,以评估外源基因的表达效果。
大肠杆菌表达系统的步骤大肠杆菌表达系统的基本步骤如下:1.选择适当的表达载体:根据需要选择合适的表达载体,包括质粒和噬菌体。
2.插入目标基因:将目标基因插入到表达载体中,通常使用限制酶切和连接酶法完成插入。
3.转化大肠杆菌:将重组载体导入大肠杆菌细胞中,通常使用热激转化或电转化的方法。
4.选择性培养:将转化后的菌液接种到选择性培养基上,以筛选含有外源基因的细菌。
5.表达蛋白质:使用适当的培养条件和诱导方法,促使含有外源基因的细菌表达目标蛋白质。
6.蛋白质纯化:利用亲和层析、离子交换层析等技术,对目标蛋白质进行纯化。
大肠杆菌表达系统的应用大肠杆菌表达系统在蛋白质表达领域具有广泛的应用。
以下是一些常见的应用领域:1.重组蛋白质的生产:大肠杆菌表达系统可用于大规模生产重组蛋白质,如重组人胰岛素等。
2.蛋白质结构和功能研究:通过大肠杆菌表达系统,可以表达和纯化具有特定结构和功能的蛋白质,用于研究其结构和功能。
3.抗原制备:大肠杆菌表达系统可以用于表达和纯化目标蛋白质,作为疫苗的抗原。
4.酶的生产:利用大肠杆菌表达系统表达酶,可以实现酶的大规模生产,用于工业生产和生物催化等领域。
总结大肠杆菌表达系统是一种高效、经济且易于操作的蛋白质表达系统。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
大肠杆菌高效表达重组蛋白策略前言重组蛋白的制备在蛋白结构分析和医疗应用领域十分重要。
药物蛋白的研究需要高纯度的重组蛋白来进行药物动力学和物理化学的研究[1]。
重组蛋白在检测酶活、连接配体、蛋白相互作用等生物学领域广泛应用。
已经表达出多种重组蛋白被证明有很大的应用潜力[2,3]。
通过基因工程改造的方法已经获得了许多性状优良的宿主菌表达系统,尤其是通过大肠杆菌可以大量表达外源基因编码的重组蛋白[4]。
但是仍然有两个问题制约着大肠杆菌表达系统对重组蛋白的表达:一个是表达量低,还有一个就是表达错误折叠的蛋白包涵体[5]。
蛋白的表达和纯化工艺一直在发展进步,但是超过30%的重组蛋白为不具有生物活性的包涵体,严重影响了重组蛋白的生产应用[6,7]。
在理想条件下,重组蛋白由强启动子进行表达,产生大量的具有生物学活性的可溶性重组蛋白。
但是,强启动子会导致重组蛋白的过表达,从而影响宿主菌体的生长并产生包涵体[8]。
在某些条件下可以通过变性、复性的方法使包涵体恢复活性[9],但是复性后的蛋白是否能够完全恢复活性仍然未可知。
一般来讲,可以通过表达条件的优化来促进蛋白的可溶性表达,比如:诱导温度、培养基组成、宿主菌的种类。
还可以通过多种方案来解决蛋白不溶的问题:蛋白重新折叠[10],构建融合蛋白[11]。
另外想要进一步增加蛋白可溶性可以与分子伴侣共表达[8]或者低温诱导[12]。
本文对目前主要的促进蛋白可溶表达的方法进行了比较全面的总结。
1.大肠杆菌表达系统的构建1.1选择表达宿主菌对于大规模的表达重组蛋白,一般选择胞表达或者周质空间表达。
与周质空间表达相比,胞表达的表达量更高,因此应用更为广泛。
在实验研究和实际生产中,已经有很多大肠杆菌表达系统广泛应用于。
在表达体系中较为常用的大肠杆菌为B菌株和K12菌株及它们的衍生菌株(表1[13])。
美国国立研究院已经认证了K12菌株的标准性以及安全的使用方案,因此K12菌株在生产应用中具有极大的优势。
但是由B菌株演变而来的BL系列菌株与K12相比,突变了lon 和ompT两个基因[14],因此具有许多表达优势:产物积累少,缺少蛋白酶,防止产物被降解。
这些优势使得BL菌株也具有非常广泛的应用[15,16,17]。
通常来讲,针对不同的重组蛋白,宿主菌的选择也是不同的。
如果重组蛋白含有大肠杆菌稀有密码子,就需要宿主能够表达针对这些密码子的tRNA,比如BL21 (DE3) CodonPlus-RIL,Rosetta (DE3)等菌株。
如果重组蛋白具有许多二硫键,则需要宿主表达环境为氧化条件的。
AD494宿主菌是硫氧还蛋白突变型,可以促进二硫键的折叠。
Origami菌株为硫氧还蛋白突变和谷胱甘肽还原酶突变,进一步加强了二硫键在细胞的形成[18]。
另外一方面,如果表达的重组蛋白对于宿主菌是有毒性的,则需要表达为包涵体的形式。
表1:常用于重组蛋白表达的宿主菌及其特点1.2 质粒的设计理想的基因转录是质粒和启动子功能协同完成的。
广泛使用的质粒都是由复制子、启动子、标记位点、多克隆位点、融合蛋白联合调控进行转录的(图1[20]。
)重组蛋白的表达量与质粒的拷贝数、结构、分离稳定性有关。
如果拷贝数低形成的mRNA数量少,蛋白的表达量就会降低。
高拷贝数可以提高蛋白的表达量,但是这也会给宿主造成代谢上的负担。
拷贝数很大程度上由起始子的复制情况决定的,也体现出质粒是严密调控还是松散调控。
图1. 质粒的基本构成高拷贝数质粒在生产应用中并不令人满意。
首先,宿主为了保存高拷贝数质粒,不得不在培养基中增加筛选抗生素。
但是FDA限制临床应用产品中添加剂的含量。
其次,高拷贝数质粒存在分离不稳定性,尤其是在低抗生素的条件下[23,24]。
第三,高拷贝数质粒的表达和维持需要大量能量,不可避免的会影响宿主菌的生长和蛋白的合成。
另外一个缺点就是,大量蛋白的过表达容易产生包涵体。
利于蛋白产品高表达的大肠杆菌表达系统应该是严密调控并且高转录效率的。
但是如果目的蛋白有毒性,则会损伤宿主细菌。
另外有些特殊的宿主只能与特定的质粒组合才能获得理想的重组蛋白。
表2. 大肠杆菌表达中经常使用的启动子有多种启动子已经成功的用于表达各类重组蛋白(表2[25,26,27,17]),。
其中,lac 启动子及其衍生物,tac和trc,在研究和工业化中有着重要的应用[28,25]。
tac和trc 启动子比lac启动子更强,这三个启动子都是由IPTG进行诱导的。
IPTG可以抑制lac启动子的阻遏,因此重组表达的水平由培养基中IPTG的浓度进行调控。
但是,IPTG价格比较昂贵,且对许多菌株有毒性作用。
为了解决这些问题,通突变筛选的方法获得了可以通过温度的调节控制热敏启动子[29]。
从T7噬菌体中得到的T7启动子也广泛的应用于蛋白重组表达。
T7噬菌体RNA聚合酶可以识别T7启动子,DNA链的延长速率比大肠杆菌RNA聚合酶快5倍。
宿主菌染色体中含有前噬菌体(λDE3)可以编码T7噬菌体聚合酶,在T7启动子衍生物L8-UV5的调控下可以进行表达。
这种新的lac启动子减少了对于AMP的依赖,也减少了对于葡萄糖抑制的敏感性。
因此T7启动子系统被认为比lac启动子系统更为强大。
通过T7启动子系统可以获得重组目的蛋白占总蛋白含量50%,但是大多以包涵体的形式存在[23,30]。
储热调控启动子系统比如λ噬菌体pL和pR启动子,也用于生产治疗性蛋白。
质粒中含有热敏性的cI857阻遏组件,可以通过温度的变化控制pL和pR启动子。
由于温控法容易操作,所以适合于大规模的产业化应用,同时也可以减少培养基的污染。
但是温控诱导也有其局限性,由于热激会抑制菌体的生长,对重组蛋白的表达也会造成一定压力。
但是对于蛋白产品的质量和产量,这些压力都是可以忽略的。
λ噬菌体pL和pR启动子也可以在低温下进行调控,应用于表达可溶蛋白或者热敏感蛋白[31]。
应用营养调控启动子phoA和trp时,培养基的营养成分要确保菌体的正常生长并且能够利用磷酸盐进行诱导。
这种诱导方法与其他的方法相比,诱导时间围非常广泛[32]。
另外一种严谨型启动子lux,从费时弧菌中分离得到,具有群体感应元件,在基因表达中由luxI进行调控,通过添加自诱导AHL物进行转录的激活。
AHL的浓度通常为IPTG浓度的1/1000,因此这种方法更加经济利于生产放大[33]。
2.融合标签的选择通过选择蛋白表达环境的方法不一定能够使蛋白可溶,因此进一步采用融合标签的方法加强可溶蛋白的表达。
在重组蛋白中增加融合标签可以增加蛋白的可溶性,并且使亲和纯化的过程更为简便[34]。
尽管这些标签最开始是用于促进目的蛋白的分选及纯化,近些年的研究表明,融合标签也可以促进蛋白的产量,加强蛋白的溶解性,并且促进蛋白的正确折叠[35]。
许多大肠杆菌表达质粒都可以在不同的启动子调控下表达多种融合标签:多聚组氨酸、麦芽糖连接蛋白(MBP)、谷胱甘肽转移酶(GST) [36]。
选择标签的主要依据是蛋白本身的特性以及蛋白处理的阶段,比如:是否为治疗蛋白,层析分离的成本以及过程可控性的影响[37]。
组氨酸标签时目前应用最为广泛的融合标签。
通过金属离子螯合层析的方法可以对组氨酸标签标记的目的蛋白进行分离纯化。
蛋白上的组氨酸标签与固载的金属离子奥何物相互作用[38]。
另外通过MBP标记的蛋白可以通过形成交联直链淀粉进行一步纯化[39],之后用含有10 mM麦芽糖的非变性缓冲液洗脱连接上的蛋白。
GST也是一个亲和力标签,通过固载谷胱甘肽可以对GST标记的可溶蛋白进行分离。
洗脱缓冲液一般含有还原型谷胱甘肽。
GST标记的重组蛋白也可以通过酶催化或者免疫测定的方法进行定量检测[34]。
想要系统的分析融合标签对于可溶蛋白的作用还是比较困难的,蛋白加入不同的融合标签后的反应也不相同[6,40]。
标签的选择十分重要,标签可能会影响天然蛋白之间的相互作用、翻译后修饰、是否可溶、重组蛋白的表达胞定位等多个方面[41,42]。
一些标签在很多条件下(含有盐离子、还原剂、表面活性剂)都可以实现功能,因此在之后的研究中可以弹性的选择不同的缓冲液组成,与标签的功能相适应[34]。
3.提高蛋白转录效率蛋白在大肠杆菌中的转录过程可以被分为4个部分:起始、延伸、终止、核糖体循环。
在大多数情况下,翻译的起始是蛋白合成决定速率的关键步骤。
这个效率由每条mRNA5’末端的翻译起始区(TIR)的序列和结构决定[43]。
翻译起始区(TIR)由四部分组成:(1)核糖体结合位点(SD)序列,(2)起始密码子,(3)核糖体结合位点和起始密码子之间的区域,(4) 在SD序列上游或是起始密码子下游的翻译增强子。
调控TIR序列可以降低或者提高大肠杆菌中重组蛋白的表达[44,45]。
已经证明了六个核苷酸的SD序列(AGGAGG)比其他更长或者更短的序列在表达绿色荧光蛋白(rGFP)方面效率更高[46]。
在这个研究中,A/U合并的加强子使rGFP 的表达增强了13倍。
在对TIR区域进行修饰的时候要避免mRNA的二级结构覆盖SD序列或者起始密码子。
在SD区域中进行单点突变会使RNA噬菌体MS2外壳蛋白的表达降低500倍[47]。
将起始密码子AUG从碱基配对的mRNA结构中暴露出来,可以有效提高大肠杆菌中IL-10的表达量,比野生型的表达量高10倍[48]。
最近,科学家研究出了翻译起始的生物学模型,可以通过设计核糖体的结合位点来改变翻译起始速率。
这个技术可以实现速率控制,并且细微精确的调整重组蛋白的表达[43]。
表达量的精确程度对蛋白的分泌十分重要,如果表达速率太快会影响细菌的分泌过程,导致最终蛋白产量低。
但是为了重组蛋白产量的最大化,翻译水平通常选择高分泌水平的表达方案[49,44]。
4.提高mRNA的稳定性mRNA的稳定性(半衰期)也会影响大肠杆菌中的表达速率。
mRNA在大肠杆菌中的降解主要是由于RNA酶的存在,包括切酶(RNase E, K and III和外切酶(RNase II 和多核酸磷酸化酶)。
RNA酶和活性和菌体的生长环境有关[50]。
对于高表达系统而言,提高mRNA的稳定性主要有两个策略:通过宿主的基因改造或者改变菌体生长条件,使mRNA的两端更具有防护性,避免被RNA酶水解[51]。
实验证明,在序列5’端非编码区中添加ompA基因可以形成一个茎环结构,有效的延长外源mRNA的半衰期[52]。
在人类IL-2的cDNA翻译终止子的位置添加来源于苏云金杆菌的penP基因,可以加强mRNA的稳定性,提高IL-2在大肠杆菌中的表达量[52]。
研究表明,在C末端添加的rne基因,可以编码RNA酶,导致mRNA的降解,因此对rne基因进行突变可以提高mRNA的稳定性,促进重组蛋白的表达量[53]。