我国单殖吸虫分布
外来鱼类单殖吸虫研究Ⅰ.食蚊鱼寄生百慕大伴盐虫Salsuginus bermudae首次在我国检出

有 关食 蚊 鱼 寄生虫 的研 究报 道 仅见 台湾棘 带 吸虫 囊 蚴和 头槽 绦虫 IJ单 殖 吸虫 未见报 道 _ . 3, 4 在光 学 显 微 镜下 对标 本 进行 了观 察测 量 ( 量度 单位 以 m计 , 括 号 中 的数 字 为 变 化 范 围 ) 标 本 保 存 于华 南 师 范 , 大 学 生命科 学 学 院鱼类 寄 生虫研 究 室.
摘 要 : 录 采 自广 东广 州 食 蚊 鱼 G mbs fn 鳃 上 的 1 单 殖 吸 虫— — 百 慕 大 伴 盐 虫 S l gnsbr u a ad& 记 a ui a i af s i 种 as iu em d eR n u
Wi s18 . l ,9 7 所获标本 与 R n e ad& Wi s的原 始描述 基本一致 , l e 但在量度 、 边缘小钩数量虫 之 锚首 虫 科 A crcp a d e 伴 盐 虫 属 n yoe hl a , i S l gn sB v r y—B r n 18 的百 慕 大 伴 盐 虫 as iu eel u e ut ,9 4 o S l gn sbr d eR n as iu e u mu a a d& Wi s 1 8 . l ,9 7 目前 , 内 e 国
~
1 形 态 描 述
宿 主 : 蚊鱼 G m ui fns a da dG r d 食 a b s a i B i n i r . af r i a 寄生 部位 : . 鳃 采集 地 点及 时间 : 广东 广州 , 0 9—1 . 20 0
1 ) 宽 2 ( 5~3 ) 卵 模 周 围环 绕 着 梅 氏腺 . 8 , 11 0 , 子
宫壁 薄 , 接 通 向 生 殖 孔 . 殖 孔 位 于肠 叉 之 后 中 直 生
斑点叉尾鮰寄生单殖吸虫中国一新纪录属(锚首虫科)

2 . 2 普 氏趋 触 虫 L i g i c t a l u r i d u s p r i c e i( Mu e l l e r , 1 9 3 6)B e v e r l e y—B u r t o n , 1 9 8 4中国新纪 录
原产 于北美 洲 , 是一 种大 型淡水鱼类 ,具有 食性 杂、 生长快 、 适应性广 、 抗病力强 、 肉质上乘等优点. 我国 1 9 8 4 年 由湖北省从美国引进原种进行驯化 , 于
1 9 8 7年人 工繁 育成 功之 后 , 在很 短 的时 间 内即推 广 到全国2 0多 个省 、 市、 自治 区进 行 养殖 .目前 , 在 湖
斑点叉尾鲴( 1 c t a l u r u s p u n c t a t u s ) 俗名沟鲶 、 美洲
鲶, 属 于鲶形 目( S i l u r i f o r me s ) 、 鲴科 ( I c t a l u r i d a e ) ,
2 结 果 与 分 析
2 . 1 趋 触 虫属 L i g i c t a l u r i d u s B e v e r l e y—B u r t o n , 1 9 8 4中国新纪 录
华 南师 范大学学报 (自然科 学版 )
2 0 1 3年 3 月
Ma r .2 01 3 J OURNAL OF S OUTH CHI NA NORMAL UNI VERS Ⅱ_ Y
第4 5卷 第2期
V0 1 . 45 No . 2
( N A T U R AL S C I E N C E E D I T I O N)
我国单殖吸虫分布

我国单殖吸虫分布:在中国,自尹文英和Sproston于1948年在华东地区首次进行调查后,从50年代至70年代,对各地单殖吸虫进行较为系统的调查的有郎所、陈启鎏、张剑英、中国科学院水生生物研究所鱼病室及苏联学者等.他们的调查结果已由纪国良等人[1]作了初步汇总.80年代后,又有郎所[2~4]、张剑英[5~7]、王伟俊[8~10],马成伦[11]、陈致和[12]、吴宝华[13]、黄少涛[14]、金燮理[15]、梁瑞刚[16]等报道了全国各地单殖吸虫的调查情况.此外还有不少零星的调查报告和新种或新记录的报道.纪国良等[1]在1982年列举中国的海水和淡水中的单殖吸虫种类总共达376种,此后十多年来,除了有大量新种报道之外,原有的单殖吸虫种类又增添了不少新的宿主记录和地理记录.因此笔者重新整理了资料,对中国内陆水体单殖吸虫种类组成进行了总结.根据笔者的统计,截至1997年7月止,中国内陆水体中已发现的单殖吸虫种类达46属575种,分别隶属于锚首虫科、指环虫科、八铗虫科、双沟盘虫科、双身虫科、三代虫科、钩铗虫科、多盘虫科和四钩虫科.各科属中种类分布数目见表1.指环虫科在内陆水体单殖吸虫中占绝对优势的地位,多达360种,占单殖吸虫总种数的62.61%;锚首虫科次之,共计111种,占19.30%;三代虫科有39种,占6.78%;双身虫科36种,占6.26%;其他5科共29种,占5.05%.锚首虫科虽包含17个属,但数量最多的锚首虫属和似鲶盘虫属所占的比例很大,两者合计77种,占全部锚首虫的69.37%.在指环虫科中,绝大部分种类属于指环虫属,达341种,占94.72%.三代虫科仅发现了三代虫属和薄片吸虫属两个属的虫种,除薄片吸虫属有1个种外,其余38个种均属于三代虫属.中国的双身虫科的单殖吸虫经姜乃澄等人[17]整理,分归于真双身虫属、前侧孔虫属、副双身虫属和华双身虫属这4个属中,其中以副双身虫属的种类占优势,为27种,所占比例为75%.八铗虫科、双沟盘虫科、钩铗虫科、多盘虫科和四钩虫科均只有少数的种属报道. 自1982年以来所报道的新种与新记录达199种之多(表2),这一方面说明科研工作者对单殖吸虫进行了卓有成效的调查与研究,同时也反映出中国内陆水体中单殖吸虫资源是相当丰富的.新种与新记录大多分布在锚首虫科和指环虫科中,两科分别为50种和120种;双身虫科中又发现了23个新种与新记录,而三代虫科中仅发现1个新种,显然这与过去对双身虫研究较少有关.此外,还有铗钩虫科1个种,两栖类中的多盘虫科也发现4个新种.没有科的新记录.八铗虫科、双沟盘虫科和四钩虫科均没有新种和新记录报道,这可能与其种类较少、宿主谱较窄有关,但也可能是由于宿主分布地区不广,缺少调查的缘故.新种和新记录的发现多在云南、广西、四川等地,而在江河平原和东北地区较少,这反映了过去对全国各地调查研究的不均衡.考虑到新疆、西藏、内蒙这些在地理条件上很有特殊性的地区还缺少调查,可以预计将来还会有较多的新种和新记录报道.在Bychowsky [1]的分类系统中,单殖吸虫(Monogenean)隶属于扁形动物门的单殖吸虫纲(Monogenoidea);Yamaguti [2]则将其定为单殖吸虫目(MonogeneaCarus,1863),细分成单后盘亚目(Monopisthocotylea Odhner,1912)和多后盘亚目(Polyopisthocotylea Odhner,1912)两个亚目;在陈心陶等编著的《中国动物志》中,单殖吸虫和复殖吸虫(Digenea)分别作为亚纲归入吸虫纲;而Boeger &Kritsky [3,4]又把单殖吸虫纲分为多钩(Polyonchoinea Bychowsky,1937)、寡钩(Oligochoinea Bychowsky,1937)和多盘(Polystomatoinea Lebedev,1986)三个亚纲。
中国淡水鱼类寄生单殖吸虫锚首虫四新种:指环虫目:锚首虫科

中国淡水鱼类寄生单殖吸虫锚首虫四新种:指环虫目:锚首虫
科
吴宝华;王淑霞
【期刊名称】《动物分类学报》
【年(卷),期】1989(014)003
【摘要】本文报道并叙述我国四种寄生于淡水鱼上的锚首虫,它们是:福建锚首虫、江西锚首虫、鄱阳锚首虫和钱塘锚首虫,分别寄生于鳅、(鱼勾)和(鱼骨)属淡水鱼的鳃丝上,分布在我国福建建瓯、江西鄱阳湖、浙江曹娥江、钱塘江等淡水江湖中。
文内各新种和相似种列表并描述了在形态特征上的相互差別,以确定该新种的成立。
【总页数】7页(P262-268)
【作者】吴宝华;王淑霞
【作者单位】不详;不详
【正文语种】中文
【中图分类】Q959.153.1
【相关文献】
1.云南省的单殖吸虫寄生于(鱼兆)科鱼类的贝氏属一新种(指环虫目:锚首虫科) [J], 李涛;毕廷菊;字应伟;骆永德
2.云南省的单殖吸虫寄生于褶鮡类的双盾新贝氏虫新种(指环虫目:锚首虫科) [J], 字应伟;骆永德;郎所
3.中国外来鱼类寄生古氏虫属(单殖吸虫,锚首虫科)3个种的描述 [J], 李海燕
4.海洋馆观赏鱼类寄生单殖吸虫四新种(指环虫目:锚首虫科) [J], 曾伯平;刘琳;等
5.华南淡水鱼类单殖吸虫的研究:ⅩⅥ.锚首虫属一新种(指环虫目:锚… [J], 李桂峰;张剑英
因版权原因,仅展示原文概要,查看原文内容请购买。
额尔齐斯河高体雅罗鱼单殖吸虫中国二新纪录种

2 结Байду номын сангаас果
21 小刺 指环 虫 . mirc nh s N b l ,1 3 ) ca a t u y e n 9 7,中国 i
分布于欧洲 、 亚洲西北部地区, 我国仅见于额尔齐斯河及 其附属水体【。20 3 09年 4月调查额尔齐斯河鱼类寄生虫 】
区系时,在高体雅罗鱼鳃上采得指环虫,经鉴定属于指
中图分 类号 : 5 .5 . Q9 91 31
文 献标识 码: A
文 章编号 : 0 03 0 (0 10 -7 30 1 0 .2 72 1 )40 1 .4
额尔齐斯河(ri Rvr Egs i ) e 发源于中蒙边境, 地处欧亚
大 陆腹 部 ,上 游 在 我 国境 内,位 于 新疆 阿尔 泰 地 区东 北
l 材料与方法
宿主高体雅 罗鱼采 自额 尔齐斯河 哈巴河 县边境河
长 006 . 2 . —0 3 。中央大钩常型, 2 0 大钩全长 O 4 . 4 . —00 , 0 4
基 部 长 00 2 .3 ,钩 尖 长 0O 8 00 2 .3 —00 4 .0 — . 1 ,内突 长
环 虫科 ( atl yia) D c o r e指环 虫 属 (a toyu) 小刺 指 yg d D c l rs的 y g
新纪 录( 1。 图 )
宿主及寄生部位: 高体雅罗鱼 L i s i au, 。 .d n es 鳃 uL n
采 集 地点 :额尔 齐 斯河 哈 巴河 县 边境 河 段 (75 ,, 4。9 N
段 。将宿 主鳃 片取下 后, 加生 理盐 水置 于玻片 上, 解 滴 在 剖 镜下 挑取虫 体,用 7% 精 固定 , 体经 洋红染 色,系 5酒 虫 列 酒 精脱 水 , 甲苯 透 明和 中性树 胶 封片 后 保存 于新疆 二 农 业 大学 动 物 医学 学 院寄 生 虫标 本 室 ; 分 虫体 经 布 氏 部 胶 固定 封 片后 ,在 显微 镜 下 测量 其 几 丁 质结 构 ,绘 制特
中国外来鱼类寄生强壮伞片虫(单殖吸虫

轻刮下虫体和黏液 , 在解剖镜下挑取虫体, 经布氏胶 固定 封片 后 , Oy p sB 5 在 lm u X一 1显微 镜下绘 制 形态
特征 图并测 量其 相关 的结 构 . 中量 度 为 : 量 度 文 mm. 为平 均值 , 括号 内为测量 的 范 围. 本保 存在 广州 大 标
华南师范大学学报 (自然科 学版 )
21 0 1年 8月
Aug 2 . 011 J OURNAL OF S OUT CHI NORMAL UNI H NA VERSTY I
2 1 年 第 3期 01
N . 2 1 o 3. 0 1
( A U ALS I N EE IIN) N T R C E C DTO
20 0 9年 8月 1 8日.
B e e。9 9中 国新 纪 录 ogr 18
() 1 属征
虫 体 中小 型 , 吸 器 染率与感染强度 : 共调查 1 4尾鱼, 中 9 其 条呈阳性 , 感染率为 6 .% , 43 最大感染强度为 40 8。
中央大 钩和 7对形 态 大 小 相 似 的 边缘 小 钩 . 背腹 具
联结片 , 其中, 腹联结片前部边缘有 2 个伞形 的膜状
结构 或腔 . 支简 单 , 肠 明显分 开 , 合 于生殖 腺后 端 , 汇 无支囊 . 殖腺 重 叠 , 于 2分 支肠 之 内. 丸 背 对 生 位 睾
着卵 巢 , 精管 环绕 左肠 支 , 输 膨大形 成贮 精囊 , 或 2 1
学生命 科 学学 院.
S h edr 强 壮 伞 片 虫 S i ie hu i ti u c n ie. c dc i rm p hm m a lt h
中国海洋鱼类单殖吸虫研究ⅩⅪ.拟八铗虫科一新记录

收 稿 日期 : 0 0— 8—0 21 0 5
基金项 目:国家 自然科学基金项 目(0 7 22) 广东省 自然科学基金项 目(3 5 8 3502 ; 0 10 )
通 讯作 者 ,igj sn .d .n dnx@ cu eu a
14 0
华 南 师 范 大 学 学 报 ( 然 科 学 版) 自
文章编 号 : 00— 4 3 2 1 ) 1 0 0 0 10 5 6 (0 1 0 — 13— 2
中国海 洋鱼 类单殖 吸虫研 究 X I X .拟八 铗 虫科一新记 录
孟杨杨 , 向英 , 熊 丁雪娟
( 华南师范大学生命科学学 院, 广东广州 5 03 ) 1 6 1
摘要: 记述 采 自广 东汕尾长体 圆够 D cpeu m c sm l kr15 ea t s ar o aBe e ,8 1鳃上 的 1种单殖吸虫—— 拟八铗虫属 P ed d— r o e s oi u cdp oaY maui与 16 l ohr a gt i , 95之 圆碜拟八铗 虫 P ed d l ohr eat u a aui16 . s oii p oadcpe sY m gt,9 5 所获标本 与 Y m gt 的原 u cd r a aui 始描述基本 一致 , 在吸铗形态 、 但 两侧 吸铗及 卵的大小上略有差异. 拟八铗 虫属及 圆够拟八铗 虫均为 中国新记 录, 长
图 1 圆Hale Waihona Puke 够 拟八 铗 虫 , 国新 记 录 中
F g r P e d dc io h r e a tr s Y ma u i 1 6 . e . iu e 1 s u o il p oa d c p eu a g t, 9 5 n r c d i h n n C ia
中国海洋鱼类单殖吸虫研究XX. 格氏虫属——新记录
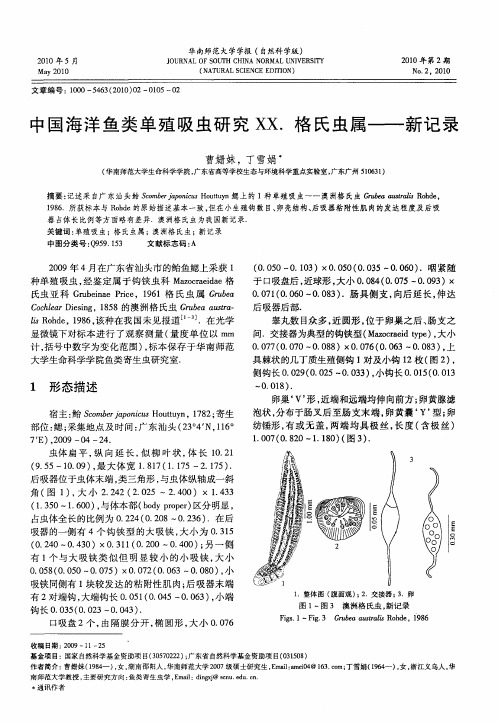
1 形 态描 述
宿 主 : So bra o i s uty ,18 ; 鲐 cm ejp nc t n 7 2 寄生 u Ho u 部位 : ; 鳃 采集 地 点及 时 间 : 东 汕 头 ( 3 4 N,1 。 广 2 。 1 6
7E ,0 9— 4— 4 )2 0 0 2 .
摘要 : 记述采 自广 东 汕头鲐 So brao i sH uty cm ejp nc ot n鳃上 的 1种单殖吸 虫一一 澳洲格 氏虫 Gu e ut l od , u u rbaas aiR he ra
18 .所获标本 与 R h e的原始描述基本 一致, 96 od 但在小生殖钩数 目、 卵壳结构、 吸器粘 附性肌 肉的发 达程度及后 吸 后
器 占体长 比例等方面略有差异.澳洲格 氏虫为我国新记 录. 关键词 : 单殖吸虫;格 氏虫属 ;澳洲格 氏虫;新记录 中图分类号 :9 9 13 Q 5 .5 文献标志码 : A
20 0 9年 4月 在广 东省 汕头市 的鲐 鱼鳃 上采 获 1
种单 殖 吸 虫 , 鉴 定 属 于 钩铗 虫 科 Mao r ia 经 zca d e格 e
有 1 与 大 吸 铗 类 似 但 明 显 较 小 的小 吸 铗 , 小 个 大
2
008 000~ .7 )× .7 (.6 .5 (.5 005 00 2 003~000 ,J . 8 ) / 、 吸铗 同侧有 i块较 发 达 的粘 附性 肌 肉 ; 吸 器末 端 后 有 2对 端钩 , 端钩 长 00 1 005~ .6 )小 端 大 .5 (.4 0 03 ,
(.5 0 0 0—0 13 0 0 0 0 0 5—0 0 0 .咽 紧 随 . 0 )× . 5 ( . 3 .6 )
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
我国单殖吸虫分布:在中国,自尹文英和Sproston于1948年在华东地区首次进行调查后,从50年代至70年代,对各地单殖吸虫进行较为系统的调查的有郎所、陈启鎏、张剑英、中国科学院水生生物研究所鱼病室及苏联学者等.他们的调查结果已由纪国良等人[1]作了初步汇总.80年代后,又有郎所[2~4]、张剑英[5~7]、王伟俊[8~10],马成伦[11]、陈致和[12]、吴宝华[13]、黄少涛[14]、金燮理[15]、梁瑞刚[16]等报道了全国各地单殖吸虫的调查情况.此外还有不少零星的调查报告和新种或新记录的报道.纪国良等[1]在1982年列举中国的海水和淡水中的单殖吸虫种类总共达376种,此后十多年来,除了有大量新种报道之外,原有的单殖吸虫种类又增添了不少新的宿主记录和地理记录.因此笔者重新整理了资料,对中国内陆水体单殖吸虫种类组成进行了总结.根据笔者的统计,截至1997年7月止,中国内陆水体中已发现的单殖吸虫种类达46属575种,分别隶属于锚首虫科、指环虫科、八铗虫科、双沟盘虫科、双身虫科、三代虫科、钩铗虫科、多盘虫科和四钩虫科.各科属中种类分布数目见表1.指环虫科在内陆水体单殖吸虫中占绝对优势的地位,多达360种,占单殖吸虫总种数的62.61%;锚首虫科次之,共计111种,占19.30%;三代虫科有39种,占6.78%;双身虫科36种,占6.26%;其他5科共29种,占5.05%.锚首虫科虽包含17个属,但数量最多的锚首虫属和似鲶盘虫属所占的比例很大,两者合计77种,占全部锚首虫的69.37%.在指环虫科中,绝大部分种类属于指环虫属,达341种,占94.72%.三代虫科仅发现了三代虫属和薄片吸虫属两个属的虫种,除薄片吸虫属有1个种外,其余38个种均属于三代虫属.中国的双身虫科的单殖吸虫经姜乃澄等人[17]整理,分归于真双身虫属、前侧孔虫属、副双身虫属和华双身虫属这4个属中,其中以副双身虫属的种类占优势,为27种,所占比例为75%.八铗虫科、双沟盘虫科、钩铗虫科、多盘虫科和四钩虫科均只有少数的种属报道. 自1982年以来所报道的新种与新记录达199种之多(表2),这一方面说明科研工作者对单殖吸虫进行了卓有成效的调查与研究,同时也反映出中国内陆水体中单殖吸虫资源是相当丰富的.新种与新记录大多分布在锚首虫科和指环虫科中,两科分别为50种和120种;双身虫科中又发现了23个新种与新记录,而三代虫科中仅发现1个新种,显然这与过去对双身虫研究较少有关.此外,还有铗钩虫科1个种,两栖类中的多盘虫科也发现4个新种.没有科的新记录.八铗虫科、双沟盘虫科和四钩虫科均没有新种和新记录报道,这可能与其种类较少、宿主谱较窄有关,但也可能是由于宿主分布地区不广,缺少调查的缘故.新种和新记录的发现多在云南、广西、四川等地,而在江河平原和东北地区较少,这反映了过去对全国各地调查研究的不均衡.考虑到新疆、西藏、内蒙这些在地理条件上很有特殊性的地区还缺少调查,可以预计将来还会有较多的新种和新记录报道.在Bychowsky [1]的分类系统中,单殖吸虫(Monogenean)隶属于扁形动物门的单殖吸虫纲(Monogenoidea);Yamaguti [2]则将其定为单殖吸虫目(MonogeneaCarus,1863),细分成单后盘亚目(Monopisthocotylea Odhner,1912)和多后盘亚目(Polyopisthocotylea Odhner,1912)两个亚目;在陈心陶等编著的《中国动物志》中,单殖吸虫和复殖吸虫(Digenea)分别作为亚纲归入吸虫纲;而Boeger &Kritsky [3,4]又把单殖吸虫纲分为多钩(Polyonchoinea Bychowsky,1937)、寡钩(Oligochoinea Bychowsky,1937)和多盘(Polystomatoinea Lebedev,1986)三个亚纲。
因此,在较高阶元的分类系统问题上,对于单殖吸虫的分类地位,一直以来各研究者的意见都有较大分歧[5]。
单殖吸虫是雌雄同体的寄生蠕虫,多寄生于水生和两栖脊椎动物上,有时候也出现于水生无脊椎动物上(如甲壳动物、软体动物等)。
单殖吸虫通常寄生于鱼鳃,有时见于宿主的皮肤、口咽腔或者与外部直接或间接连通的其它器官,例如鼻孔、眼、耳朵、泻殖腔、直肠腺和膀胱等,甚至偶尔能寄生于鱼的血液系统。
不同于复殖吸虫的是,单殖吸虫直接发生,没有中间宿主,他们的幼体从卵中孵化出来,逐渐发育成成体。
然而,也有例外,一些海水鱼的单殖吸虫如Pricea 和Gotocotyla 能寄生在多种鱼类的鳃,它们并不直接发育成熟,而是在这些鱼被“终末”宿主(大型鲭鱼)捕食后才发育成熟[6,7]。
据记载,人类对单殖吸虫的最早研究是1776 年Muller 在丹麦的Hippoglossusvulgaris 鱼体上发现,当时以为是一种水蛭,并以Hirudo hippoglossi 命名;1818年,Blaninvelle 将之命名为内吸虫Entobdella hippoglossi。
直到20 世纪六十年代以前,人们对单殖吸虫的研究集中于形态与分类学以及生活史。
六十年代以后,Kearn [8,9]对鳎内吸虫Entobdella soleae 进行了详细深入的研究,表明人们对单殖吸虫的研究已深入到了生理学的层次。
自七十年代以来,学者们以流行病学方法为寄生虫-宿主种群建立大量的数学模型,并对寄生虫生态行为展开了全面的实验研究[10]。
我国单殖吸虫的研究始于上世纪四十年代。
1948 年,尹文英和Sproston [11]报道了采自华东地区沈家门鲬鱼鳃上的锚首虫及海盘虫各一种。
在海洋单殖吸虫方面的研究,黄宗国[12]和申纪伟等[13]分别报道了黄渤海物种海域的15 种和13 种单殖吸虫。
近年来,张剑英等[14]对我国南海鱼类单殖吸虫做了大量的研究,其编著的《中国海洋鱼类单殖吸虫》描述单殖吸虫200 多种。
随着新技术的不断应用及学科之间的相互渗透,单殖吸虫的研究内容也逐渐扩展,主要涉及生态学、动物地理学、形态学、组织学、组织化学与超微结构、区系分类学、方法学和病害防治等各方面。
单殖吸虫病的危害性寄生虫是指寄居在其它生物体内或体外的生物体,它们部分或全部地从宿主获得营养,并普遍地表现出一定程度的适应模式,且至少潜在地对宿主产生一定的伤害。
一些理化因子例如温度对寄生虫有重要的影响,但是与非寄生物种相比,环境因子所起的作用更直接和连续,影响更大。
寄生虫的环境条件(例如:温度、PH、氧气等)是宿主,它的食物是宿主的食物或是宿主体自身,它的生境是宿主体,并且它的转移依赖于宿主。
寄生虫病可能对宿主的体重或繁殖产生重大的影响,可能改变宿主的种群特征,因此,对寄生虫病进行研究有重要的经济意义[6]。
大部分单殖吸虫是鱼类的体外寄生虫,并频繁发现于海洋鱼类。
它们一般寄生在鱼的鳃、皮肤、鳍,但有一些物种能寄生在鱼的直肠、尿管、体腔、甚至血液系统。
单殖吸虫一般是通过小钩、吸铗、吸盘、摩擦垫子、表面的棘、粘着腺,或者这些结构的组合来附着在鱼的体表。
单殖吸虫通常以其后吸器上之钩插入寄生部位,造成寄主部位的病变产生不良的后果,导致病鱼幼鱼的死亡;或破坏器官完整性,引起其他病原生物的入侵,造成炎症,产生病变;或吮吸鱼血、黏液、刺激宿主产生大量的分泌物,破坏正常的生理活动。
淡水鱼类指环虫病病理学的研究开展的较早,各家研究结果基本一致:指环虫的致病性及损伤程度决定于宿主的年龄和寄生虫的数量和大小,鱼类鳃指环虫病都可以引起鳃瓣缺损、出血、组织增生和坏死[15,16,17,18,19]。
李立伟[20]在2002 年研究了寄生在花鲈上的菇茎指环虫和逆转指环虫,研究8虫群落,发现在污染的湖中寄生虫群落的平均密度和多样性都是最高的。
Morand等[52]的研究表明:物种丰富度似乎更多地归因于宿主的特征而非寄生虫的作用,指出就单殖吸虫而言,当控制潜在的偏差(如系统发生的影响等),宿主大小是物种丰富度的主要决定因子。
Lo 等[48]对法属波利尼西亚岛的三种珊瑚礁鱼类,黑真雀鲷Stegastes nigricans,三带圆雀鲷Dascyllus aruanus 和斑点九棘鲈Cephalopholis argus 的寄生虫群落进行了调查,发现这三种鱼的体外寄生虫平均密度与宿主体长也有正相关关系;因而认为外寄生虫群落似乎受生物因子影响更大(例如宿主免疫等)。
Rohde 等[53]调查了来自澳大利亚大堡礁的长吻裸颊鲷Lethrinus miniatus 后生类外寄生虫的群落生态学,检获12 种单殖吸虫,6 种桡足类,2 种成体单足类和几种幼体单足类,1 种蛭,认为,群落较高的物种丰富度可能与宿主的栖境相关。
Sasal 和Morand [54]以地中海海水鱼类的单殖吸虫种类物种作为例子,使用独立对照的方法来控制取样和系统发育的可能带来的混杂影响,研究了几个影响寄生虫物种丰富度的因子,他们的结果表明,宿主的大小是影响单殖吸虫物种丰富度和专一性的主要因子。
李敏敏[55对头鲻的寄生虫进行了研究,认为寄生于头鲻的寄生虫种类较多是由于宿主不做长距离的洄游。
单殖吸虫种群动态学寄生虫种群(Population)是由占有一定时空领域的单一虫种的所有个体组成。
寄生虫种群(Population )分为内种群(Infrapopulation )和总种群(Suprapopulation),前者为在一定时间内一种寄生虫在单个宿主体内的所有个体数量,后者为在一定时空里一种寄生虫在所有宿主的各个发育阶段的所有个体的数量,这里的前提“一定时空”对Margolis 等[56]的“某一生态系统”作了修订[44]。
关于海水鱼类寄生虫的生态研究还很少,并且大多数都是来自低温海区的,这些地区由于不同季节温差较大,水温成了寄生虫季节动态变化的主要因子,其它非生物因子(如盐度)的影响难以显现出来,对于全面了解单殖吸虫的种群生态学显然是不全面的。
国内关于单殖吸虫的研究大多局限于区系调查和物种描述上,生态方面的研究很少。
淡水单殖吸虫的生态研究主要有:聂品[57]对养殖鳜鱼Siniperca chuaisi 鳃上7和未成熟阶段的异钩盘虫附着在鳃丝上,并集中在前一对鳃弓的腹面,随后渗透到皮下组织。
Whittington 等[43]研究了单殖吸虫的前吸附区与宿主特异性的关系,指出其在识别宿主表皮的粘液中起重要作用。
单殖吸虫生态学单殖吸虫种群生态学是研究单殖吸虫种群的数量、分布以及单殖吸虫种群与非生物因素和其它生物种群相互关系的科学,如:季节动态、空间分布、中间关系、单殖吸虫种群和宿主种群之间的关系、同种单殖吸虫在不同地区、不通宿主和不同季节其可测部分(硬质结构)的变化等[12]。