基因沉默
基因沉默的研究及应用实例简介

用 RNaseIII 消化长片断 siRNA表 双链RNA制 siRNA表 达载体 备siRNA 达框架
体外转录
• 以DNA Oligo为模板,通过体外转录合成siRNAs, 成本相对化学合成法而言比较低,而且能够比化 学合成法更快的得到siRNAs。不足之处是实验的 规模受到限制。 • 值得一提的是体外转录得到的siRNAs毒性小,稳 定性好,效率高,只需要化学合成的siRNA量的 1/10就可以达到化学合成siRNA所能达到的效果, 从而使转染效率更高。 • 最适用于:筛选siRNAs,特别是需要制备多种 siRNAs,化学合成的价格成为障碍时。 • 不适用于:实验需要大量的,一个特定的siRNA。 长期研究
负对照
(1)一个完整的siRNA实验应该有负对照。 (2)作为负对照的siRNA应该和选中的siRNA序列有 相同的组成,但是和mRNA没有明显的同源性。 ( 3 )通常的做法是将选中的 siRNA 序列打乱,同样 要检查结果以保证它和其他基因没有同源性。
制备siRNAs的方法
化学 合成 体外 转录
策略
• 利用RNAi降解CYP17mRNA,使CYP17基因所编码蛋白 的合成明显降低,降低CYP17基因蛋白对其下游基 因的调控,在很大程度上阻断了肿瘤细胞的应答, 建立一种基因修饰前列腺癌治疗策略,提高前列腺 癌的治疗效果。
新鲜前列 腺癌标本
组织中总 RNA的提取
反转录
siRNA的 制备
siRNA
目录
1 2
基因沉默的概述 RNA干扰
3
4
基因沉默的应用实例
RNAi的生物学意义
1.基因沉默
• 定义:基因沉默(gene silencing)是指 生物体中特定基因由于种种原因不表达或 者是表达减少的现象。
RNA干扰与基因沉默

RNA干扰与基因沉默RNA干扰(RNA interference, RNAi)是一种通过特定RNA分子介导的基因沉默机制,被广泛应用于生物学研究、基因治疗等领域。
本文将深入探讨RNA干扰的原理、应用及未来发展方向。
一、RNA干扰的原理RNA干扰是一种高度保守且广泛存在于真核生物中的生物学过程。
它主要通过三种类型的RNA分子实现基因沉默:microRNA (miRNA)、small interfering RNA(siRNA)和Piwi-interacting RNA (piRNA)。
其中,siRNA是最为常见和被广泛应用的一种。
在RNA干扰中,siRNA与RNA诱导酶复合物结合形成RNA诱导沉默复合物(RISC),RISC会通过碱基互补的方式与靶向RNA结合,并介导靶向RNA的降解,从而达到沉默该基因的效果。
这一过程使得基因的转录和翻译被有效地抑制,实现基因的沉默。
二、RNA干扰的应用1. 基因功能研究:RNA干扰技术被广泛应用于基因功能的研究中。
通过设计特定的siRNA,可以实现对目标基因的沉默,从而观察基因沉默对生物体的生理和生化过程产生的影响,揭示基因在细胞和生物体中的作用机制。
2. 疾病治疗:RNA干扰技术在基因治疗领域具有巨大潜力。
通过设计特异性的siRNA,可以实现对致病基因的沉默,从而治疗遗传性疾病、肿瘤和病毒感染等多种疾病。
此外,RNA干扰还可以用于研发新型药物和治疗手段。
3. 植物保护:在植物领域,RNA干扰技术也被广泛应用于植物保护。
通过设计特定的siRNA,可以实现对害虫和病原菌基因的沉默,从而提高作物的抗病虫性,减少对农药的依赖,实现绿色农业的发展。
三、RNA干扰的未来发展方向随着RNA干扰技术的不断发展,未来有望在以下几个方面取得重要进展:1. 靶向性增强:未来的RNA干扰技术将更加注重提高siRNA的靶向性,减少对非靶向基因的影响,从而提高沉黙效率和生物安全性。
2. 交叉学科应用:RNA干扰技术将与生物信息学、纳米技术等学科相结合,开拓全新的应用领域,如基因组编辑、精准医学等。
基因沉默方法汇总

基因沉默⽅法汇总从遗传学中⼼法则看,基因表达沉默⽆⾮从两个⽔平研究,先将具体⽅法总结如下1基因组DNA⽔平1.1 构建基因重组打靶载体基因功能研究中出现最早最成熟的技术就是基因敲除. 他是根据同源重组的原理, 利⽤分⼦⽣物学技术增强和减弱甚⾄灭活某特定靶基因表达⽔平, 然后观察实验动物整体功能状态的变化, 推测靶基因的功能[1]. 基因敲除技术可在整体动物⽔平研究基因功能, 但是完全基因敲除使⼩⿏所有细胞基因组上都存在基因的缺失或突变, 有些重要的靶基因敲除或导⼊会严重影响动物胚胎的发育, 导致胚胎早期死亡或严重的发育缺陷, 使得突变⽆法传代, 不利于在⼩⿏各个发育阶段进⾏该基因功能的分析. 条件基因敲除技术(conditional gene knock out)的建⽴为此难题找到了解决的办法. 1994年, Gu et al [2]应⽤Cre/loxP重组酶系统实现了外源基因的时间特异性表达, 在此基础上条件基因敲除技术得以形成. 条件基因敲除技术是在基因敲除基础上结合Cre/loxP系统⽽形成的, 他可以做到在特定时间, 组织, 细胞中将靶基因敲除, 从⽽可以真实的反映特定组织或细胞中靶基因被敲除或修饰后的结果, 避免在发育早期所有细胞和组织中完全敲除⽬的基因后可能产⽣的胚胎早期死亡或严重的发育障碍[3-5]. 基因敲除的优点是基因灭活效果确切可靠, 缺点是技术复杂, 费时费⼒.1.2 反义寡核苷酸技术反义寡核苷酸技术是指采⽤⼀类经⼈⼯合成或构建的反义表达载体表达的寡核苷酸⽚段, 长度多为15-30个核苷酸, 导⼊细胞或者个体体内, 根据碱基互补原理, 通过与靶DNA或者mRNA结合形成双链杂交体激活核酸酶H, 裂解靶mRNA阻断蛋⽩质的翻译, 或者与DNA结合成三链结构或与单链DNA结合成双链结构以阻⽌靶基因的复制或转录, 以及与mRNA AP位点结合⼲扰其剪接、加⼯和运输, 在mRNA⽔平上发挥作⽤, 从⽽⼲扰其表达, 阻⽌其翻译成蛋⽩质[6]. 具体的作⽤机制⽬前尚未完全清楚. 反义寡核苷酸因为是针对特定的靶mRNA(DNA)的序列设计合成, 因此具有极⾼的特异性, 并且容易设计和体外⼤量合成. 另外反义寡核苷酸不含病毒序列, 不会产⽣免疫反应, 也不会整合⼊宿主染⾊体内, 这都为其作为药物应⽤于临床提供了可能. 1998年, 第1个反义药物Vitravene(Fomivirsen)被美国FDA批准通过, ⽤以治疗由巨细胞病毒(cytomegalovirus)引起的艾滋患者的视⽹膜炎[7]. ⽬前反义寡核苷酸技术在动物体内外的应⽤已经⾮常普遍[8-9]. 今后要注意的是, 如何对反义寡核苷酸进⾏更加有效的化学修饰以提⾼其稳定性, 延长其半衰期, 增加其作⽤时间和如何定点作⽤于特定部位, 使靶组织最⼤效率地吸收反义核酸, 提⾼其作⽤效果, 减轻毒副作⽤[10].1.3 甲基化寡核苷酸技术表观遗传学(epigenetics)是与遗传学(genetics)相对应的概念. 遗传学是指基于基因序列改变所致基因表达⽔平变化, 包括基因突变、基因杂合丢失和微卫星不稳定性等;⽽表观遗传学则是指基于⾮基因序列改变所致基因表达⽔平变化, 包括DNA甲基化、组蛋⽩脱⼄酰化和染⾊质构象变化等; 表观基因组学(epigenomics)则是在基因组⽔平上对表观遗传学改变的研究, 以抑癌基因为代表的CpG岛甲基化所致基因转录失活已经成为肿瘤表观基因组学研究的重点内容[23-25]. 所谓DNA甲基化是指在DNA甲基化转移酶的作⽤下, 基因组5'端CpG⼆核苷酸的胞嘧啶第5位碳原⼦上共价结合⼀个甲基基团. 由于DNA甲基化与⼈类发育和肿瘤疾病的密切关系, 特别是CpG岛甲基化所致抑癌基因转录失活, DNA甲基化已经成为表观遗传学和表观基因组学的重要研究内容, 研究DNA甲基化与肿瘤的关系成为当前分⼦⽣物学的热点之⼀. ⽬前的研究已经表明: 肿瘤细胞和组织中存在异常的DNA甲基化状态, 表现为基因组整体甲基化⽔平降低, 导致遗传不稳定性增加;组织特异性基因的启动⼦区域出现从头甲基化从⽽导致基因被关闭; 原癌基因多为低甲基化或不充分甲基化, 低甲基化使原癌基因活化, 导致重新开放或异常表达, 形成突变热点, 增加染⾊体的不稳定性; 抑癌基因多为过度甲基化, 过度甲基化导致表达失活[26-27]. 这些因素综合起来导致基因表达异常, 引起细胞恶变, 最终导致肿瘤的发⽣[28-30].在哺乳动物中, 甲基化仅影响DNA链上鸟嘌呤前的胞嘧啶(CpG). 通常细胞中CpG⼆核苷酸的甲基化分布并不是均⼀的, ⼤约50%的基因在启动⼦区域有CpG⼆核苷酸的富集现象, ⼀般该区域的长度从0.5-2 kb不等. 该区域与基因的转录有密切的关系, 通常处于⾮甲基化状态. 处于⾮甲基化状态的启动⼦, 环绕的染⾊质呈现为开放的构象, 允许转录因⼦和其他的激活物靠近. 此外, 转录因⼦的占据也使其他的转录抑制因⼦和染⾊质重塑蛋⽩等难以接近启动⼦, 最终表现为启动基因表达[31-33]. 相反, CpG岛⾼甲基化的启动⼦则呈现为关闭的构象, 不但使转录因⼦⽆法靠近, ⽽且还有助于甲基胞嘧啶结合蛋⽩﹑转录辅阻遏蛋⽩﹑DNA甲基转移酶等对转录有抑制作⽤的蛋⽩结合于启动⼦区, 启动⼦失去功能, 结果基因转录灭活⽽沉默[34-35]. 很多资料表明, 基因启动⼦异常⾼甲基化可以导致其转录灭活[36-39]. Zhu et al [40]通过甲基化寡核苷酸诱导ERβ基因启动⼦区和外显⼦区CpG岛特异性甲基化发现, 在前列腺癌细胞中, 是ERβ基因启动⼦区(⽽不是外显⼦区)CpG岛的⾼甲基化导致了ERβ基因转录失活. 去甲基化试剂作⽤前列腺癌细胞后, ERβ mRNA恢复表达.甲基化寡核苷酸技术是利⽤针对靶基因合成的甲基化寡核苷酸⽚段(methylated oligonucleotides,MON)与基因的其中⼀条链互补结合形成半甲基化DNA. 半甲基化DNA表现为复制叉样结构, 为DNA甲基化转移酶1(DNA methyltransferase-1, DNMT1)的优先底物, DNMT1使第1链迅速甲基化. MON与结合点分离, 甲基化的第⼀链与未甲基化的互补链退⽕形成第2个半甲基化DNA底物, 同样表现为复制叉样结构, 为DNMT1作⽤的优先底物. 结果两条链均发⽣甲基化, 随后, 同样的道理, 甲基化扩布到邻近的CpG⼆核苷酸中, 最后整个基因的靶位点(如启动⼦)发⽣完全甲基化[41]. 如上所述, 基因启动⼦甲基化可以导致其转录失活, 因此通过甲基化寡核苷酸技术, 我们就可以对所要研究的⽬的基因进⾏特异的灭活[42-43]. 此外, 尚有资料表明,CpG岛甲基化表型(CIMP)能够改变染⾊体的构象, 产⽣微卫星不稳定性(MSI)现象, 引起靶基因突变失活[44-49]. 这种有趣的现象使遗传学和表观遗传学之间建⽴起了桥梁, 为研究两者之间的联系提供了思路[50]. 基因CpG岛的这种甲基化修饰具有可遗传性, 能够对发育、⽣理、环境、病理等不同的信号作出反映, 使遗传信息的表达按⼀定程序发⽣变化, 参与完成细胞的时空调控和适应调控, 在胚胎发育障碍等先天性疾病以及恶性肿瘤发病机制中起⾄关重要的作⽤.甲基化寡核苷酸技术的实施路线包括: (1)MON的设计与合成: MON是包含20余个碱基的⼀⼩段寡核苷酸⽚段, 与⽬的基因启动⼦区对应位置的碱基序列完全相同, 不同的是MON⽚段中的5'端CpG⼆核苷酸中的胞嘧啶环5位碳原⼦发⽣甲基化(m5CpG). ⽽GenBank数据库中原始的⽬的基因启动⼦区的CpG⼆核苷酸并未发⽣甲基化修饰(CpG). MON⽚段设计时应注意⾄少包含3个CpG⼆核苷酸, 理论上MON⽚段中所含的m5CpG越多越好, 这样才能达到更好的甲基化诱导效果. 为了防⽌在细胞中被酶降解, 寡核苷酸⽚段还需进⾏硫代磷酸化等修饰, 此外还可以标记荧光来⽰踪,同时还需要设计⾮甲基化寡核苷酸⽚段(unmethylatedoligonucleotides, UMON)作为对照. UMON碱基序列与MON完全相同, 不过没有进⾏CpG ⼆核苷酸甲基化修饰. MON的合成与引物合成⼀样, 可由DNA⾃动合成仪来完成, 价格⽐较低廉. MON的作⽤: 针对体外细胞的应⽤, 通常采⽤的是脂质体介导的基因转染, 体内实验的资料⽬前不多. ⽬前把MON导⼊体内外的基因转染⽅法主要有两⼤类: ⼀类为病毒介导法, 即利⽤去掉了致病基因的病毒序列作为载体, 将外源靶基因导⼊靶细胞内, 常⽤的有逆转录病毒、疱疹和腺病毒等改建的病毒载体; 另⼀类为⾮病毒介导法, 包括物理法(显微注射、⽓溶胶、基因枪和缝线等)、化学法(磷酸钙沉淀法、脂质体法和葡聚糖法等)和⽣物学法(细胞融合法和受体法等). (3)效果评价: 提取基因组DNA和总RNA以及蛋⽩质, 采⽤甲基化特异性PCR(MSP)、亚硫酸氢盐测序(BSP)、RT-PCR、Western blot等⼿段进⾏相关的分析. 其中, MSP法可以了解是否发⽣了特异性的甲基化诱导,BSP可以对那些发⽣了甲基化的CpG进⾏精确定位[51-53]. RT-PCR、Western blot则可以从不同分⼦⽔平判断基因灭活的效果及其他相关分析.⽬前甲基化寡核苷酸技术的应⽤多集中在肿瘤细胞系[60-61], 这实际上就是把他作为⼀种基因沉默的⼯具应⽤到过度表达的肿瘤相关基因的研究中, 借此观察这些靶基因的功能. 甲基化寡核苷酸技术在此所起的作⽤可以⽤前述的其他基因沉默⼯具代替. 然⽽, 甲基化寡核苷酸技术尚具有基因敲除等技术所不能代替的优点. 例如, 针对肿瘤细胞中抑癌基因启动⼦发⽣甲基化⽽失活, 我们就可以将正常的组织细胞或动物模型作为研究对象, 采⽤甲基化寡核苷酸技术来诱导其发⽣甲基化⽽失活, 模拟肿瘤细胞和组织中该基因的甲基化⾏为, 从⽽⽣动地再现肿瘤细胞和组织中抑癌基因甲基化灭活的过程, 达到相关研究的⽬的. 这主要是基于甲基化寡核苷酸技术的作⽤机制和肿瘤细胞中的抑癌基因的基因型. 实际上肿瘤相关基因启动⼦⾼甲基化导致转录沉默, 甚⾄发⽣MSI突变⽅⾯的⽂献数不胜数[44-49]. 因此, 甲基化寡核苷酸技术的应⽤⼤有前景, ⽽这正是其他基因沉默⼯具所不能⽐拟的. ⽬前甲基化寡核苷酸技术应⽤于体内实验的资料不多, 毒副作⽤的研究很少, 和反义寡核苷酸技术以及⼲扰RNA技术等其他基因治疗⼿段存在的问题⼀样. 如何⾼效、靶向、安全地把这种神奇的甲基化寡核苷酸⽚段应⽤到个体体内是⽬前存在的问题之⼀,这有赖于靶向载体研究的进展.1.4 锌指核酸酶技术 锌指核糖核酸酶(ZFN)由⼀个 DNA 识别域和⼀个⾮特异性核酸内切酶构成。
基因沉默研究进展

基因沉默研究进展摘要:基因沉默(gene silencing)是指生物体中特定基因由于种种原因不表达或者表达减少的现象。
基因沉默是基因表达调控的一种重要方式 ,是生物体在基因调控水平上的一种自我保护机制 ,在外源 DNA 侵入、病毒侵染和DNA 转座、重排中有普遍性。
对基因沉默进行深入研究 ,可帮助人们进一步揭示生物体基因遗传表达调控的本质 ,在基因工程中克服基因沉默现象 ,从而使外源基因能更好的按照人们的需要进行有效表达;利用基因沉默在基因治疗中有效抑制有害基因的表达 ,达到治疗疾病的目的 ,所以研究基因沉默具有极其重要的理论和实践意义[1]。
关键词:基因沉默,转录水平基因沉默,转录后水平基因沉默,病毒介导的基因沉默.基因沉默(gene silencing)是指生物体中特定基因由于种种原因不表达。
一方面,基因沉默是遗传修饰生物(genetically modified organisms )实用化和商品化的巨大障碍 ,另一方面 ,基因沉默是植物抗病毒的一个本能反应 ,为用抗病毒基因植物工程育种提供了具有较大潜在实用价值的策略—RNA介导的病毒抗性(RNA-mediated virus resistance ,RMVR)[2~4]。
基因沉默现象首先在转基因植物中发现,接着在线虫、真菌、水螅、果蝇以及哺乳动物中陆续发现。
基因沉默主要发生在两种情况,一种是转录水平上的基因沉默(transcriptional gene silencing, TGS) ,另一种是转录后基因沉默(post- transcriptional gene silencing, PTGS)。
RNA干扰(RNA interference, RNAi)是近几年发展起来的转录后基因阻断技术,RNAi在2002年被Science评为全球十大科技突破之一,作为一种在细胞水平的基因敲除工具,RNAi 正在功能基因组学领域掀起一场革命[5]。
基因沉默
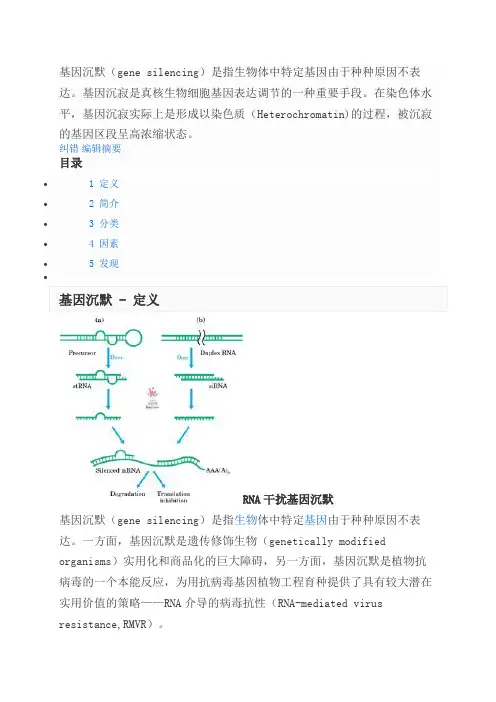
RNA干扰基因沉默基因沉默(gene silencing)是指生物体中特定基因由于种种原因不表达。
一方面,基因沉默是遗传修饰生物(genetically modified organisms)实用化和商品化的巨大障碍,另一方面,基因沉默是植物抗病毒的一个本能反应,为用抗病毒基因植物工程育种提供了具有较大潜在实用价值的策略——RNA介导的病毒抗性(RNA-mediated virus resistance,RMVR)。
转基因植物和转基因动物中往往会遇到这样的情况,外源基因存在于生物体内,并未丢失或损伤,但该基因不表达或表达量极低,这种现象称为基因沉默。
转基因沉默分为转录水平的沉默(TGS)和转录后水平的沉默(PTGS)。
TGS是指转基因在细胞核内RNA合成受到了阻止导致基因沉默,PTGS是指RNAi——基因沉默指南基因沉寂(Gene Silencing) 也可以被称为“基因沉默”。
基因沉寂是真核生物细胞基因表达调节的一种重要手段。
在染色体水平,基因沉寂实际上是形成以染色质(Heterochromatin)的过程,被沉寂的基因区段呈高浓缩状态。
基因沉寂需要经历不同的反应过程才能实现,包括组蛋白N端结构域的赖氨酸残基的去乙酰基化加工、甲基化修饰(由甲基转移酶催化,修饰可以是一价、二价和三价甲基化修饰,后者又被称为'过度’甲基化修饰(Hypermethylation) ) 、以及和甲基化修饰的组蛋白结合的蛋白质(MBP)形成“异染色质”,在上述过程中,除了部分组蛋白的N端尾部结构域需要去乙酰化、甲基化修饰之外,有时也许要在其他的组蛋白N端尾部结构域的赖氨酸或精氨酸残基上相应地进行乙酰化修饰,尽管各种修饰的最终结果会导致相应区段的基因“沉寂”失去转录活性。
这个“原则”就是目前尚没有真正完全清楚的“组蛋白密码”(Histone Code)。
能够与甲基化组蛋白结合的蛋白质有sir1/2/3/4,这是一组被称为"Silencing Informative Repressor"的蛋白,其中,Sir2就是上文中的“去乙酰化”酶,而Sir1/3/4则负责与甲基化修饰的组蛋白结合"沉寂”相应的染色质为异染色质。
基因沉默名词解释

基因沉默名词解释基因沉默,指的是抑制或抑制正常的基因功能。
基因沉默可以在多种水平上发生,从分子层次到细胞层次,从细胞层次到组织层次,再到整个机体组织水平。
基因沉默可分为三类,即转录抑制、调节抑制和调节转录抑制(TGS)。
转录抑制是指基因转录过程中的抑制,它是由转移因子介导的,通常是由抑制基因的非编码RNA、DNA复合物或其他蛋白质抑制有效的HTR导致的。
当转录因子在基因上聚集时,它们可以抑制此基因上的有效拷贝数量及其表达,从而降低或抑制基因的功能。
调节抑制是指在基因转录后的调控过程中,由抑制蛋白质通过影响mRNA或蛋白质的稳定性来抑制基因表达。
调节抑制可以在不同水平上发挥作用,例如在细胞中可以抑制mRNA和蛋白质的形成,在组织水平上可以抑制蛋白质的稳定性和细胞分化,从而抑制基因表达。
这种抑制机制可以使基因表达更加精细,可以更好地调节基因功能,从而调节机体的新陈代谢。
调节转录抑制也叫TGS,它是一种可以在基因组织水平上实现基因沉默的技术,它可以实现非编码RNA涉及的基因表达调控。
在基因水平上,TGS可以改变mRNA和蛋白质的形成方式和稳定性,从而抑制基因表达,在组织水平上,TGS可以影响细胞分化,从而抑制机体的器官及组织的新陈代谢。
此外,TGS还可以通过调控细胞的基因表达,影响细胞的生长、分化和功能,从而抑制疾病发病。
基因沉默在生物的发育过程中具有重要作用,它可以控制基因的表达,从而调节细胞的发育和机体的新陈代谢。
目前,基因沉默技术被用于各种疾病治疗,如癌症、心脏疾病和神经系统损伤等,这些技术可以改变基因表达水平,从而抑制疾病发病。
未来,基因沉默技术可能在生物医学领域展开广泛的应用,例如可以用于器官的再生、药物的研发等。
同时,基因沉默技术在生物安全性、社会安全性和科学道德上也可能引起讨论,因此,在基因沉默技术的应用时,还需要综合考虑法律、人文、社会等因素。
基因沉默是一种重要的基因调控技术,它可以影响基因表达、影响细胞发育和机体新陈代谢,还可以用于疾病治疗,因此,基因沉默技术将在未来发挥更多重要的作用。
引起基因沉默的原因
引起基因沉默的原因研究表明,引起基因沉默的原因很多,转基因的拷贝数和构型、在植物上的整合位点、转基因的转录水平等都与沉默有关,外界环境如过高的温度、过强的光照也会增加基因沉默发生的几率和产生时间,此外,外源基因的表达还受植物发育因子(如亲本年龄)的影响。
因此,植物转基因沉默的作用机制可能不是单一的,而是各种机制共同作用的结果,是植物本身的防御系统和外界环境因素协同作用的产物。
转基因沉默可以发生在染色体DNA水平、转录水平和转录后水平三种不同的层次上。
1.染色体DNA水平的转基因沉默发生在染色体DNA水平的转基因沉默叫做位置效应(positioneffect)。
当导入的外源基因随机地插入到宿主基因组时,如果被导入到转录活跃区,就有可能进行高水平的转录,如果外源基因插入转录不活跃区,则只能进行低水平的转录或不能转录。
按照染色质高级结构组织的环状结构模型,核基质结合区(matrixattachmentregions,MARs)作为边界元件与核基质结合,使两个MAR之间的基因片段被界定成一个独立的染色质环(1oop),并作为隔离子(insulator)阻挡邻近染色质区的顺式调控元件对环内基因的影响,使位于染色体环内的基因可作为一独立的表达调控单位而存在。
MAR可能使转基因在受体基因组整合后形成独立的环状结构,从而提高转基因的表达水平并减少转基因在不同株系表达差异2.转录水平的基因沉默发生在转录水平上的转基因沉默叫做转录失活。
反向重复的基因或转基因可以进行异位配对,配对的DNA作为信号,使DNA异染色质化或从头甲基化,这样转录过程就会受到抑制。
此外,DNA-RNA协同(association)也是造成转录水平基因沉默的原因之一。
(1)转移基因及其启动子甲基化甲基化是活细胞中最常见的一种DNA其价修饰形式,它通常发生在DNA的GC和CN G序列的C碱基上,C甲基化的频率在哺乳动物及高等植物中部比较高。
甲基化修饰在基因表达、植物细胞分化以及系统发育中起着重要的调节作用。
RNA基因沉默的原理及应用
RNA基因沉默的原理及应用1. 概述RNA基因沉默是一种常见的基因表达调控机制,它通过特定的RNA分子干扰来抑制基因的表达。
这一机制在生物学研究和疾病治疗等领域具有重要的应用价值。
本文将重点介绍RNA基因沉默的原理及其在基础研究和生物医学领域的应用。
2. RNA基因沉默的原理RNA基因沉默主要包括两种机制:小干扰RNA(small interfering RNA,siRNA)和microRNA(miRNA)。
2.1 小干扰RNA(siRNA)siRNA是由双链RNA或先导RNA(precursor RNA,pre-RNA)在细胞内被特定核酸酶剪切而产生的小分子RNA。
siRNA通过结合到特定的mRNA上,形成RNA诱导沉默复合物(RNA-induced silencing complex,RISC),RISC可以帮助降解mRNA,从而阻止其转录和翻译过程。
siRNA可以通过人工合成和转染等方式引入细胞内,用于研究基因功能和疾病治疗等领域。
2.2 微小RNA(miRNA)miRNA是一类由内源性基因转录产生的短小非编码RNA分子。
miRNA作为一种调控因子,可以与目标mRNA结合并抑制其翻译过程,从而影响基因表达。
miRNA在细胞分化、发育和疾病进程中发挥重要作用。
研究发现,miRNA与多种疾病的发生和发展密切相关,如癌症、心血管疾病等。
因此,miRNA具有潜在的临床应用价值。
3. RNA基因沉默的应用RNA基因沉默技术在基础研究和生物医学领域有广泛应用,下面将介绍其主要的应用方面。
3.1 基因功能研究通过沉默特定基因的表达,可以研究该基因在生物体内的功能。
siRNA或miRNA可以通过转染或基因敲除等方式引入细胞中,从而实现对目标基因的沉默。
这种方法广泛应用于生物学研究领域,帮助科学家了解基因的功能和调控机制。
3.2 疾病治疗RNA基因沉默技术在疾病治疗中有着巨大的潜力。
通过沉默特定基因的表达,可以干扰疾病相关基因的功能,从而达到治疗的目的。
基因沉默
Hale Waihona Puke 共抑制• 共抑制现象普遍存在于转基因植物中。 共抑制的发生是由于外源基因编码区与 受体细胞基因间存在同源性而导致外源 基因与内源基因的表达同时受到抑制。
基因压制
• Cogoni在粗糙脉抱霉的转化实验中发现, 外源基因可以抑制自身和相应内源基因 的表达,他们把这一现象定为基因压制。 他们发现,基因压制是可逆的,而且这 种逆转会伴随有外源拷贝的丢失。基因 压制发生在转录后水平,导致已复制的 稳定态mRNA大量减少。
转录水平上的基因沉默
• 转录水平基因沉默是指对基因专一的细 胞核RNA合成的失活,它的发生主要是 由于基因无法被顺利转录成相应的RNA 而导致基因沉默。
引起转录水平基因沉默的 机制主要有以下几种:
• • • • • .基因及其启动子甲基化 基因及其启动子甲基化 同源基因间的反式失活 后成修饰作用 重复序列 位置效应
2 基因沉默的和分类及机制
• 根据作用部位和作用机理的不同,将基因 沉默分为转录水平上的基因沉默 (Transcriptional gene silen-cing,TGS) 和转录后基因沉默(Post transcriptional gene silencing,PTGS),TGS发生在细胞 核内,一般由于启动子失活而导致基因转 录无法正常进行,而PTGS则是指基因能 正常转录
转录后基因沉默
• 指基因在细胞核内能稳定转录,但在细 胞质里却无相应稳定态mRNA 存在的现 象。与转录水平基因沉默不同,转录后 水平基因沉默具有逆转性,即受抑制基 因通过减数分裂可以恢复表达活性,表 现为减数分裂不可遗传性。
已发现主要有以下几种转 录后水平基因沉默现象
• 共抑制 • 基因压制 • RNA 干扰
基因沉默技术:治疗遗传性疾病的新希望
基因沉默技术:治疗遗传性疾病的新希望在医学的广阔天空中,基因沉默技术如同一颗新星,其光芒正在逐渐照亮治疗遗传性疾病的未来。
这项技术,被科学家们形象地比喻为“分子剪刀”,能够精确地剪除或修复导致疾病的基因序列,从而为患者带来新的希望。
首先,让我们来了解什么是基因沉默技术。
简单来说,它是一种通过特定的分子机制,使特定基因的表达受到抑制或完全停止的技术。
这就像是给有问题的基因按下了“静音键”,阻止它继续制造有害的蛋白。
那么,这项技术为何能成为治疗遗传性疾病的新希望呢?让我们来看几个例子。
首先,对于一些由单一基因突变引起的疾病,如囊性纤维化、镰状细胞贫血等,基因沉默技术可以直接针对这些突变基因进行干预,从根本上解决问题。
其次,对于一些复杂的多基因疾病,如癌症、心脏病等,虽然不能直接治愈,但基因沉默技术可以作为一种辅助治疗手段,帮助减轻症状或延缓病程。
当然,任何新技术的出现都会伴随着挑战和争议。
基因沉默技术的安全问题是人们关注的焦点之一。
毕竟,我们是在对生命的基本单位——基因进行操作,稍有不慎就可能引发严重的后果。
此外,如何确保技术的准确性和有效性,也是科学家们需要不断探索的问题。
尽管如此,我仍然对基因沉默技术抱有乐观的态度。
我相信,随着科学研究的深入和技术的进步,这些问题最终将得到解决。
而且,与其他治疗方法相比,基因沉默技术具有不可比拟的优势。
它不仅可以针对病因进行治疗,还可以实现个性化医疗,根据每个患者的具体情况制定治疗方案。
在我看来,基因沉默技术就像是一把双刃剑。
一方面,它为我们提供了治疗遗传性疾病的新途径;另一方面,它也带来了新的挑战和风险。
我们需要谨慎而明智地使用这项技术,确保它真正造福于人类。
最后,我想用一个比喻来结束这篇文章:基因沉默技术就像是一艘航船,正驶向未知的海域。
虽然前方可能充满了风浪和暗礁,但只要我们坚定信念、勇往直前,就一定能够到达理想的彼岸。
在这个过程中,科学家、医生和患者都需要携手合作,共同面对挑战、分享成果。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
基因沉默摘要随着基因技术的迅速发展和广泛应用,在转基因技术实践中首先暴露出来的外源基因不能按照预期设想进行表达的问题越来越显得普遍,而人们对基因沉默现象的不断深入研究和探索,不仅揭示出了基因沉默的发生机制,也在一定程度上推动了新技术的产生和应用,这不仅推动了基因研究领域的发展,更在遗传群体构建、疾病治疗等方面建立了新方法、新体系,为生物学技术的发展做出了贡献。
关键字基因沉默分类机理应用1.引言基因沉默(Gene Silencing),又称为基因沉寂,是真核生物细胞基因表达调节过程中的一种特殊生理现象,是指细胞基因在表达过程中受到各种因素的综合作用而导致基因部分区段发生“沉寂”现象,从而失去转录活性并不予表达或表达减少。
该现象最先于1986年Peerbolte在转基因植物研究中所发现,随后科学家在线虫、真菌、水螅、果蝇以及哺乳动物中陆续发现了基因沉默现象的存在。
转基因沉默是基因沉默现象最为频发和常见的,这也是转基因为何在受体难以百分之百全部表达的因素之一,其基本特征是导入并整合到受体基因组的外源基因在当代或后代中表达活性受到抑制。
研究发现,其主要原因是由于转基因之间或转基因与内源基因之间存在着序列同源性,因此转基因沉默又被称为同源性依赖的基因沉默(homology-dependent gene silencing)。
根据沃森-克里克的核酸碱基互补配对模型,基因沉默可能涉及到DNA-DNA、DNA-RNA以及RNA-RNA三种不同形式的核酸分子之间的互作,简单地说就是插入的外源DNA或自身基因区段在核内高浓度的RNA作用下,能够与内源反向DNA 或者RNA进行碱基互补配对,并且在核内被重新甲基化,进而导致基因沉默;而另一种可能则是内源基因与转基因转录生成的RNA之间互补配对生成可被RNases酶性降解的双链RNA(dsRNA),其水解直接导致基因的不表达,即基因沉默效果。
从染色体水平上看,基因沉默现象的实质是形成异染色质(Heterochromation)的过程,检查发现被沉寂的基因区段往往呈现出高浓缩状态,显然,这在一定程度上也决定了被沉寂基因的难表达性。
实验早已证明,在高度浓缩的基因区段,正常的DNA转录活动是难以进行并维持的,换言之,即一旦形成异染色质进入高度浓缩状态,那么相应区段的基因片段就必然因为不能被转录而表现无活性。
然而看似简单的这一点变化,其中的分子机理却错综复杂,有些至今依旧是生物学研究中难以解答的。
事实上,由于现代转基因技术以及遗传学研究领域的飞速发展和扩张,以此为代表的基因表达高效性和特殊遗传样本的构建,基因沉默的现象及生理学意义正逐渐成为基因技术的前沿,乃至分子遗传学研究的热点。
基于此,了解和掌握相应的基因沉默机制对于每一个生物科学研究者而言都具有显而易见的优势,尤其是在转基因以及遗传学领域。
2.基因沉默的分类根据现有实验或理论表明,基因的沉默多发生在转录水平和转录后水平,据此,部分文献将基因沉默分为两类,即转录水平的基因沉默(transcription gene silencing, TGS)以及转录后水平的基因沉默(post-transcriptional gene silencing, PTGS)。
然而有些基因沉默类型显然是难以服从上述两种分类的,例如转基因沉默中的位置效应(position effect),其沉默机制简言之可以表述为转基因导入位点处于异染色质状态或外源基因与受体基因区段存在显著差异进而被细胞防御免疫系统所识别并消除,这两种情况都是发生在转录前的染色体DNA自身的变异或效应,并不具备转录水平基因沉默定义中在转录水平上发生沉寂的条件。
因此,基因沉默又可以划分为三类,除了上述两种外,还有染色体DNA水平的基因沉默。
在现有实验技术手段下,可以利用DNA免疫印迹(Northern blotting)、RNA酶保护分析(RNase protection assay)和细胞核进行转录分析(Nuclear run-on assay)对上述基因沉默层次加以区分和判断。
不过鉴于基因沉默现象用于人工操纵这一事实,基因沉默又可以根据沉默的发生是否处在人为控制或操纵之下,可以简单划分为被动基因沉默和主动基因沉默。
根据这一划分原则,目前所知的任意沉默机制均可包含其中,例如转基因技术中目的基因常由于内源性因子的干扰而不能按预期设想进行相应表达的现象归入前者,但诸如RNA干扰技术(RNAi)、基因敲除等人工干预手段导致的基因沉默则应划归为后者,即主动基因沉默。
2.1被动基因沉默在基因沉默研究最为深刻的转基因领域,在实验中常常会发生导入的目的基因在受体细胞中不能够按照实验预期进行表达,亦或是表达减少的情况,研究上可以将这种受受体细胞内源性因子影响而导致的基因沉默称为被动沉默。
实验表明,转基因沉默发生的层次水平具有多样性和特殊性,不过总结起来可以划分为染色体DNA、转录和转录后三种不同的层次。
在已知的理论或实验依据下,染色体DNA水平的基因沉默包含位置效应、DNA甲基修饰以及组蛋白乙酰化;在转录水平上,基因沉默的可能类型包含基因启动子甲基化、同源基因反义失活、后成修饰作用(epigenetic effect)以及多拷贝重复序列等;而转录后水平的基因沉默,则分为基因共抑制(co-suppression)以及基因压制。
除此之外,由生物自我保护机制引发的基因沉默也属于被动基因沉默范畴。
2.1.1染色体DNA水平外源基因所整合到的目的基因组的特性将直接决定外源基因的表达情况,这种效应称之为位置效应。
位置效应大体上可由两种情况所引发,一是目的基因整合位点或基因区段处于受体基因组中高度甲基化或异染色质化的异染色质区和重复序列区,由于这些基因区段的特殊性和高度不可表达性,由此外源基因在整合到相应区段时就不可避免地陷入沉默。
事实上,这种情况下的转基因仍是完整和可表达的,只限于所整合位点的特殊性而丧失表达的可能。
二是外源转基因在碱基组成上和整合区段有着显著的差异,进而被受体细胞的防御免疫系统所识别并做出相应反应,由此该区段被限制表达或不表达。
不过细胞防御系统究竟是如何做出反应并对相应区段限制表达的机理现在并无确凿证据说明,结合X染色体随机失活现象和基因组印记研究,酶性识别或是失活中心假说均可以在一定程度上解释并预测某些现象,但实验上还没有给出相关研究说明。
DNA甲基化修饰和组蛋白乙酰化是基因沉默在染色体DNA水平上的另一种机制。
很早以前研究就已经证明了DNA甲基化参与了染色质凝聚状态的形成,事实上,这也是最早发现的基因修饰途径之一,几乎存在于所有高等生物的基因表达调控中。
其主要作用机理是通过甲基化来改变位点DNA的构象,从而影响了蛋白质与DNA的相互作用,抑制了转录因子与启动区DNA的结合效率。
至于组蛋白乙酰化的分子机制也大体类似,即组蛋白在完成翻译后,其氨基末端常发生包括乙酰化在内的多种共价修饰,并最终相互联合或依次被特定蛋白酶或其它复合体所识别、结合,为激活或抑制相应基因转录的染色质相关蛋白提供结合位点。
2.1.2转录水平启动子甲基化是转录水平上的基因沉默中较为常见的一种,比之于上述的DNA甲基化实质上并没有区别,只是发生的区段和在基因表达的先后时间顺序上有所差异。
研究发现,转录水平上的甲基化多发生在基因5,端的启动子区域,基因的表达水平与其整合位点的甲基化程度密切相关,而这种甲基化的发生条件则依赖于一种被称之为从头甲基化转移酶(de novo DNA methytransferase, DNMT)的识别和加工。
这种基因区段性的甲基化是通过核酸碱基互补配对实现传递的,现有实验证明这种甲基化是可以由基因5,端的启动子区域延伸到基因3,端,而核苷酸配对过程得以使甲基化在基因组中自由传递,进而导致整合区域的基因均不表达或随机表达,这与基因转座子相类似,去甲基化试剂可以逆转转基因沉寂现象即可说明这一理论。
而同源基因间的反式失活是近年来新发现的一种基因沉默机制,是一种由于拥有同源序列的沉默位点和其他位点的DNA的互作而引起的基因沉默。
其作用原理是通过顺式作用甲基化失活的基因作为一种“沉默子”,并对其他与之分离且具有同源性的靶基因施加反式作用使之甲基化失活,从而达到沉默基因的目的。
后成修饰作用是一类比较特殊的基因沉默,与其他类型不同的在于后成修饰作用并不改变基因的序列或碱基组成,基因沉默的方式表现在细胞内因子对功能的即时关闭和解除的控制上。
显然,这种转基因沉默与受体植物的核型构成有关。
此外,外源基因如果通过PCR技术扩增并以多拷贝的形式整合到受体基因组DNA 的同一位点上,则导入的基因会发生自身重复序列间的相互配对,形成可被甲基化酶特异性识别的结构,进而发生甲基化作用,抑制基因的表达,由此转录失活,这种的基因沉默称为重复序列间基因沉默,其作用机理可能与异染色质化相关蛋白质结合体的识别配对序列结构有关。
但有研究认为,重复序列很可能是通过一位配对,即多拷贝重复基因序列之间以及外源基因与转录产物之间的配对,使整整合位点的染色质发生异染色质化或DNA的甲基化,在空间上形成位阻,进而阻碍了转录因子与外源基因的接触,这与实验相吻合,也是大多数研究者所共同持有的观点。
2.1.3转录后水平转录后水平的基因沉默是指基因在细胞核里能够稳定转录,但在细胞质里却无相应的稳定mRNA存在的现象,简言之即转录后沉默发生在细胞质中,转录产物能在细胞核中积累但在细胞质mRNA被迅速降解或不能正常被加工。
在植物基因研究中上述现象称为共抑制,但事实上,共抑制是最常见的转录后水平基因沉默,该现象最先由Napoli在CHS基因的矮牵牛花中发现,并普遍存在于转基因植物中。
与TGS相比,PTGS显得更为普遍和常见。
转录后水平的基因沉默是指导入的外源基因引起内源基因或外源与内源基因两者共同的沉默,本质上是由基因转录本(转录本:由一条基因通过转录形成的一种或多种可供编码蛋白质的成熟mRNA)过量引起的。
由反转录酶—聚合酶连锁反应(Reverse Transcription polymerase chain reaction, RT-PCR)实验,在出现共抑制现象的转基因受体细胞中积累着高浓度的正常mRNA,但细胞质中却几乎检测不到mRNA,由此说明这类基因沉默不是发生在转录水平,而是属于转录后的调控过程。
而在真菌领域,上述基因沉默则被称为基因压制(quelling),并由Cogoni 等人最先在粗糙链孢霉的转化实验中所发现,其特征是外源基因可以抑制自身和相应内源基因的表达。
作为转录后水平的基因沉默,该发现有力的证据是在实验中发现已获得的稳定态mRNA的大量减少。
相较于转基因植物中的共抑制,亦或是真菌中的基因压制,在动物中这类基因沉默则以RNA干扰(RNA interference,RNAi)的形式出现,但三者在本质上具有极大的相似性和可比较性,事实上,后者正作为遗传学研究中的一种常规基因操纵手段,在遗传材料制备和遗传信息改良等方面正发挥着基于以上的若干事实,人们提出了许多假设和模型试图来解释和验证共抑制现象存在的分子机理,但无论是RNA阈值模型(Dougherty和Park,1995)、异常RNA模型(Jorgensen等,1998)、RNA介导的病毒抗性模型(Mueller、Lindbo 等,1993)还是现在较为学术界所接受和认同的反义RNA模型(Lee等,1997),也只是能够在一定程度上部分解释了实验中的共抑制现象,并无法圆满阐述清楚这一基因沉默现象的本质。