补体的发现
第五章-补体

④ 肽链降解后常具有新的生物学活性或与其它补体 的降解成分重新组合,形成新的活性分子。
⑤ 性质不稳定,不耐热,56℃30分钟即可灭活, 室温下很快失去活性。
五、补体的合成与代谢 ① 合成:肝细胞,单核/巨噬细胞,造血细胞,纤维 母细胞,内皮细胞,生殖细胞,脂肪细胞,神经 细胞。
② 代谢:非常快,血浆中补体每天约有一半更新。
二、替代途径
三、 MBL途径(凝集素途径)
MBL:甘露糖结合凝集素 FCN:纤维胶原素
MBL途径是指细菌或病毒表面的甘露糖蛋白与血 清中的MBL或FCN结合,进而激活C4、C2、C3 的活化途径。
三、 MBL途径(凝集素途径)
1、激活物:病原微生物表面以甘露糖或半乳糖为末端糖基 的糖结构 2、参与成分:MBL(FCN),MASP,C2-C9 3、激活过程: 病原入侵 TNF,IL等诱导机体产生MBL等 急性期蛋白 MBL结合到细菌表面的甘露糖 激活丝氨 酸蛋白酶MASP 活化C4,C2等 形成MAC杀伤入侵的病 原微生物
导C9分子聚合,产 生穿膜孔道,引起细 胞溶解
补体经典激活途径激活过程(二)
二、替代途径
该途径越过C1、C4、C2直接激活C3,故又称C3途径或旁路 途径。 激活物:某些细菌、内毒素、酵母多糖、葡聚糖等,为补 体激活提供保护性环境和接触性的表面。 参与替代途径的激活与调节因子:B因子、D因子、P因子为 激活因子;H因子、I因子为抑制与调节因子。 在细菌感染早期,即可发挥重要的抗感染作用
4. 补体的中和及溶解病毒的作用
抗体对病毒的中和作用,阻止病毒对宿主细胞的吸附
和穿入。
只有补体也可出现溶解病毒(有囊膜)的现象,
如由补体介导引起RNA肿瘤病毒溶解的现象。所有 C型RNA病毒,均能被灵长类动物新鲜血清所溶解
第18章-补体检测及补体参与试验

系统性红斑狼疮(SLE)•病因:•遗传•内分泌(雌激素(受体),催乳素、生长激素)•感染(麻疹、副伤寒、单纯疱疹、风疹、EB病毒等)•物理因素(紫外线等)•药物•免疫异常(BC功能亢进,TC失衡,CK表达异常,淋巴细胞凋亡异常)•致病机制:•机体产生大量抗细胞核样物质(DNA、RNA、核内可溶性蛋白)的抗体,形成大量IC,沉积于周身毛细血管,关节滑膜,心脏瓣膜等处,导致全身性损伤实验室检查•1.血常规:三系减少(RBC、WBC、PLT)•2.血沉:增快•3.毛细血管镜检查•4.免疫血清学检查:•狼疮细胞、类风湿因子、补体等补体检测•1.补体是什么?•2.为何要检测补体?•3.如何测?有哪些方法?第十八章补体的检测及补体参与的试验Contents第一部分 补体第二部分 补体的检测第一节 血清总补体活性测定第二节 单个补体成分的测定第三节 补体受体的测定第三部分 补体参与的试验掌握:补体的概念及其理化性质;血清总补体活性测定(CP-CH50)的实验原理;补体结合试验的原理。
熟悉:CP-CH50方法、结果判定及方法评价;单个补体成分测定。
了解:其他第一部分 补 体补体的发现1补体的生物学特性2补体系统的激活、调控3补体的生物学功能4补体与疾病的关系5Jules Bordet (1870-1961),Discoverer of Complement ( Nobel Prize for Physiology or Medicine in 1919)19世纪末,在发现抗体后不久,Bordet 通过霍乱弧菌溶菌实验发现,新鲜血清中存在一种不耐热的成分,可辅助特异性抗体介导的溶菌作用。
Ehrlich 同时独立发现了类似现象,他认为这种因子是抗体发挥溶细胞作用的必要补充条件,故将其命名为补体一、补体的发现正常豚鼠血清(溶菌)感染霍乱弧菌的豚鼠的血清霍乱弧菌菌液(溶菌)感染霍乱弧菌的豚鼠的血清56℃30分钟(凝集)(溶菌)(凝集)Presumptions:1. There is a component in the fresh serum that helps the antibody to lyse the bacteria.2. The chemical property of this component is not stable.3. This component is not antigen specific.补体(complement,C):存在于人和动物血清、组织液和某些细胞上的一组与免疫有关,但无抗原特异性,激活后具有酶样活性的,不耐热的糖蛋白。
补体
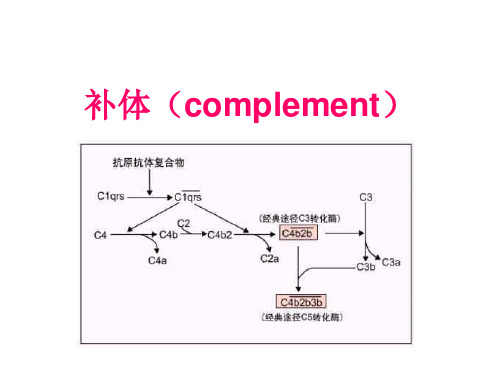
1. 补体概述
补体是存在于正常人体或脊椎动物血清与组织液中的一组与 免疫有关,经活化后具有酶活性的蛋白质 由30多种可溶性蛋白质和膜结合蛋白组成的多分子系统 补体系统正常在体内以无活性酶原形式存在,需经活化才能 发挥效应
Liver hepatocytes Lymphocytes monocytes complement proteins
补体激活的MBL途径
3.3 补体激活的旁路途径
活性物质是脂多糖、酵母多糖、葡聚糖、凝聚的IgA 和IgG4等物质 步骤包括:C3转化酶(C3bBb)的形成 C5转化酶(C3bBb3b)的形成 C3正负反馈
补体三条激活途径全过程示意图
4. 补体的生物学作用
4.1 细胞溶解作用 补体系统激活后,通过级联反应可在靶细胞表面 形成许多MAC,导致靶细胞溶解,在感染早期主要通 过旁路途径和MBL途径,抗体产生后主要通过经典途 径
4Hale Waihona Puke 2 调理作用 C3b、C4b可促进吞噬细胞的吞噬作用
图中CR1和IgFc受体可分别介导调理作用,同时参与调理作用称联合调理 作用
4.3 趋化作用 趋化因子C5a导致吞噬细胞向感染部位聚集,发生炎 症反应
5. 补体与抗体结合部位
抗原与抗体结合,抗体构象发生改变,暴露出补体结合位点 (CH2)
思考题:
1. 补体在特异性免疫中起重要作用,补体的作 用有特异性吗 2. 补体激活的三条途径,其实质有什么不同
3. 补体的激活途径
3.1补体激活的经典途径 激活物质是抗原抗体复合物 参与成分是C1-C9 激活过程是三个阶段,即识别阶段、活化阶段和膜攻击 阶段
膜攻击复合物
3.2 MBL激活途径 是指细菌和病毒表面的甘露糖残疾与血清中的MBL结合, 进而激活C4、C2、C3的活化途径 病原微生物感染早期,体内的M和中性粒细胞产生TNF 、LI-1和IL-6,诱导肝细胞合成与分泌急性期蛋白,其中与 补体激活的有C反应蛋白和甘露聚糖结合凝集素(MBL)
补体系统的基本概念

下列哪种成分无酶活性 A、C1s B、C1r
C、P因子
D、D因子
E、I因子
参与溶菌作用的补体成分是 A、C3 B、C42
C、C1
D、C56789
E、C423
补可抑制C1酯酶活性的是 A、H因子 B、I因子
C、C1INH
D、S蛋白
E、C4bp
预习提纲
1、什么叫MHC? 2、MHC与MHC分子有何联系与区别?
识别阶段
C1酯酶形成
C5b + C8 C9
C56789——MAC
C1q
C1r
C1s
C1分子模式图
C1q 与 IgM 的结合
C1q 与 IgG 的结合
C4
C4a C2 C4b
C2a
C2b
C4b
C4b2b
C3转化酶的形成
C3a C5a C3
C5
C5b
C4b2b C3b
C4b2b3b
C5转化酶的形成
MBL激活途径
4、补体系统各成分的生物学作用
补体经典激活途径中各成分的 激活顺序为 A、C123456789 B、C124536789 C、C145236789 D、C124356789 E、C142356789
补体替代激活途径中的C3转化 酶是 A、C1s B、C42 C、C423 D、D因子 E、C3bBb
红细胞
补体的免疫黏附作用
清除免疫复合物
补体的调理作用
细菌 C3b受体
吞噬细胞
补体调理杀菌作用
补 体 系 统 各 成 分 的 生 物 学 作 用
C1q——识别免疫复合物、识别病毒膜蛋白 C4a——过敏毒素 C4b——组成C3、C5转化酶、参与免疫粘附 C2b——组成C3、C5转化酶
补体(中文,本科)

活化
Mg++
C4
C4b + C2
C4b2a (C3转化 酶) C2b C3 C3b C3a C5b-C6,7,8,9
C4a
C4b2a3b
(C5转化 酶)
细胞溶解
C5 C5a
攻击
终末途径
C5b+C6+C7+C8+C9 = MACs 补体膜攻击单位结构
MACs 造成的细胞膜损伤
C6 C7 C5b C8 C9多 聚体
• 补体的分类:
补体固有成份 补体调节蛋白
C1INH、C4BP、 H、I、S蛋白和 血清羧肽酶等,
补体受体
C1~C9,
B、D、P因子
MCP, DAF, HRP
C1qR、 C3b/C4bR (CRI)、 3dR(CRII)、H因 子受体、C3a和 C5a受体等
补体固有成份的组成、命名、生成部位和理 化特征
(二) 旁路活化途径
LPS,多糖,凝聚Ig等
Ba P因子
D C3C3b+B因子C3bBb(P)
C3a 正反馈调节环路 细胞溶解
活化
C3bnBb(P)
(C3转化酶)(C5转化酶)
C5b-C6,7,8,9
C5
C5a
旁路激活途径的特点
1 C3 天然活化,LPS等多糖类物质及聚合的Ig可促进 其活化。 2 C1、C4 和C2不参与,B因子、D因子、P因子参与 3 产生不同于经典途径的C3转化酶和C5转化酶 4 C5活化后的攻膜过程与经典途径相同 5 含有一个C3活化的正反馈调节环路 6 可在机体早期抗感染免疫中起作用,也参与特异性 免疫,放大补体活化效应
4.24补体

+
+丝氨酸蛋白酶
病原体甘 露糖残基
MASP
C4b2a
C3转 化酶
C4b2a3b C5转化酶
C2 C2a+C2b
C5bC 6789
膜攻击复合物
14
补体三条激活途径
❖ 一、经典途径: ▪ 参与补体成分:C1-C9 ▪ 激活物质:抗原抗体复合物(IC)
❖ 二、旁路激活途径: ▪ 参与补体成分:C3、B、D、P因子,C5-C9 ▪ 激活物质:某些细菌内毒素和真菌等。
C1由C1q、C1r、C1s组成
3
2.补体激活的调节蛋白 主要以可溶性和膜结合 两种形式存在。前者包括C1抑制物、P因子、I 因子、H因子、C4结合蛋白、S蛋白、SP40/40 等;后者包括促衰变因子、膜辅助蛋白、同种 限制因子和膜反应溶解抑制因子等。
3. 补体受体(CR) 补体受体可与相应的补体 活性片段或调节蛋白结合,介导补体生物学效 应 。 包 括 CR1~CR5 、 C3aR 、 C2aR 、 C4aR 、 C5aR等
6
第二节 补体系统的激活
❖一、补体活化的经典途径 ❖二、补体活化的旁路途径 ❖三、MBL途径 ❖四、补体活化的共同末端效应
7
抗原抗体复合物
C1
C4
C2
经典激活途径
C4b2a
C3
C4b2a3b
C5
C5a+C5b
C67
C89
C5b6789,MEMBRANE ATTACK COMPLEX
抗原抗体复合物
24
补体的生物学作用:
❖1、溶细胞、溶菌及抗病毒作用 ❖2、调理作用 ❖3、引起炎症反应 ❖4、清除免疫复合物
25
第五节 补体与疾病的关系
第一节 补体的概述

第一节补体的概述19世纪末,Bordet发现并证实在人和动物新鲜血清中存在一种不耐热的成分,可辅助和补充特异性抗体介导的溶解细菌,称为补体(complement,C)。
现已知补体是存在于新鲜血液中的一组不耐热的成分,由30多种蛋白组成,故称为补体系统(complement system)。
补体成分广泛存在于血清、组织液和细胞表面。
通常体内大多数补体组分以无活性形式存在,只有当病原体或抗原-抗体复合物与其相遇、使之活化后才能发挥溶解细胞、介导炎症反应、调理吞噬、调节免疫应答和清除免疫复合物等生物学作用。
补体系统是一个具有精密调控机制的蛋白质反应系统。
补体系统不仅是机体固有免疫系统的重要组成部分,在特异性免疫应答过程中也发挥重要作用。
补体过度活化、补体组分缺陷和功能障碍与多种疾病的发生发展密切相关。
一、补体系统的组成和命名(一)补体系统的组成根据补体系统各成分的功能不同,可将其分为补体固有成分、补体调节蛋白和补体受体三类。
1.补体固有成分是指存在于血浆和体液中、构成补体基本组成的蛋白质,包括:①参与经典途径的C1(C1q、C1r、C1s)、C4和C2;②参与凝集素途径的甘露聚糖结合凝集素(MBL)、纤维胶原素(FCN)和MBL相关的丝氨酸蛋白酶(MASP);③参与旁路途径的D因子、B因子和备解素(properdin,P因子)等;④参与三条激活途径共同成分C3、C5、C6、C7、C8和C9。
2.补体调节蛋白是指存在于血浆中和细胞膜表面,通过调节补体激活途径中关键酶而控制补体活化强度和范围的一类蛋白质分子,包括血浆中的Cl抑制物、I因子、H因子、S蛋白、C4结合蛋白、过敏毒素灭活因子等可溶性调节蛋白和细胞膜表面的膜辅助蛋白(MCP)、衰变加速因子(DAF)、同源限制因子等膜结合蛋白。
3.补体受体(complement receptor,CR)存在于不同细胞膜表面,能与补体活性片段结合,介导多种蛋白生物学效应的分子,主要包括CR1、CR2、CR3、CR4、CR5、C3aR、C4aR、C5aR、C1qR及H因子受体(HR)等。
第五章 补体

2 补体调节蛋白
以可溶性或膜结合形式存在,具有调节和控制补体活化
作用的蛋白分子,包括C1抑制物、I 因子、C4结合蛋
白、H因子、S蛋白、促衰变因子、膜辅助蛋白、同种 限制因子等。
3 补体受体
存在于细胞表面,介导补体活性片段或调 节蛋白发挥生物效应的各种受体(CR),
如CR1~CR5、C3aR、C2aR、C4aR等。
一、补体激活的经典途径
( The Classical Pathway)
(一)经典途径的激活物及激活条件
Ag—Ab免疫复合物(immune
complexes,IC) (IgM、IgG1、IgG2、IgG3)
某些逆转录病毒的胞膜蛋白等
一个C1q分子必须同时与两个以上补体结合位点结
酸性粘多糖、肝素、鱼精蛋白、纤溶酶、组织蛋白酶、
补体系统的组成
1)固有成分:
经典及MBL途径的 前端反应成分: C1、 C4、C2、C3 MBL、 MASP
旁路途径的前端 反应成分:
C3、B、D、
2)调节成分: C1-INH、 I因子 C4bp、 H因子、 MCP、DAF、 HRF 3)补体受体:
P因子
共同末端反应成分 C5、C6、C7、C8、 C9
作用靶
C1r, C1s C3b,C4b C4b,C3b C4b C5b67
主要功能
抑制丝蛋白酶 阻止C3b与Bb结合 蛋白裂解,钝化 加速C4b2a衰变 阻止形成膜孔
整合的膜蛋白:
DAF 多数血细胞, 上皮及内皮细 胞 红细胞,淋巴 细胞、单核细 胞、嗜中性粒 细胞、血小板 C4b2a, C3bBb C8,C9 加速C3转化酶衰变
CR1、CR2、 CR3、CR4、 C3aR、C4aR
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
第四章补体
1、补体的发现,新鲜免疫血清,体内体外溶筒,称为免疫溶菌现象为60℃30分钟灭活。
2、与抗原刺激无关。
第一节补体的组成
1、三组成分 CR—Cg ①周有成分,(14分子),②激活调控分子
B,D,P,H,I因子③补体受体。
调节因子
2、表示法 C3→C3a+C3 C3bi C4
3、产生细胞肝C
μφ
肠粘膜上皮细胞
4、理化性质,均为糖蛋白,大多数为β球蛋白,少数为α,γ球蛋白。
固有成分对热不稳定,通常56℃30分被灭活。
第二节补体系统的激活
一、经典途径(Classical pathway)
激活物质IfG1,IfG2,IfG3,IfG4,分别与行形成复合。
1、识别阶段(单位)C1。
2、活化阶段(单位)C1,C2,C3。
3、膜攻击阶段(单位)C5—C9。
C5667嵌在胞膜上.
激活物,Ab IgG 1,G3,IgM。
二、替代途径(旁路激活途径)(Alternative pathiay)
1、正常生理情况下的准备阶段。
2、旁路途径的激活。
激活物:LPS,肽聚糖,病毒感染细胞,肿瘤细胞等。
3、激活效应的放大。
4、两条激活途径的比较。
三、补体激活过程的调节
(一)体液中可溶性调节分子的作用 1、自行衰变的调节。
Czb ,Csb
2、体液中灭活物质的调节。
(1)C 1,抑制物
可与C 1不可逆地结合,使C 1失去酯酶活性。
(2)C 4结合蛋白(C4binding protech C4bp )能竞争性地抑制C 4b 与C 2b 结合。
(3)I 因子,又称C 3b 灭活因子(C3binactiuator )裂解C 3B →C 3vI 。
(4)H 因子(Factor H )
能竞争性地抑制B 因子与C 3B 的结合,还能使C 3B 从C 3bBb 中置换出来。
(5)S 蛋白(S protein ) 能干扰C 15667与的胎膜结合。
(6)C 8结合蛋白。
可阻止C 5678中的C 8与C 9的结合。
3、同种限制因子(homslogous restiction factor,HRF ) 又称Cg 结合蛋白,存在于正常人红细胞,单核细胞淋巴细胞及血小板上,主要作用是通过对C 5b678复合物中C 8分子的结合,阻断C 9与C 8的结合及C 9分子的聚合,使自身细胞膜上不能形成由C 5b6789膜攻击复合867。
(二)膜结合性调分子的作用
广泛存在于血细胞和其它组织细胞表面,其主要功能是保护宿主自身组织细胞免遭补体介导的破坏作用。
1、膜辅因子蛋白(membrane cofoc protacn MCP ) 是一种穿膜蛋白(CD46),可与一些组织细胞表面粘附的C 4b / C 3b 结合,主要作用是协助I 因子裂解灭活自身组织细胞表面结合的C 4b / C 3b ,从而抑制C 3转化酶的形成,病原微生物和其它旁路途径激活物表面缺乏MCP ,因而粘附在他们表面的C ab / C 3b 可保持活化,并易与C 2/B 因子结合。
2、促衰变因子(decay accelerting factor.DAF ) 为单链腹蛋白分子(CD55),分布在一些组织细胞上,而病原微生物和某些补体激活物表面缺乏,DAF 能与上述细胞表面粘附的C 4b /A
C 3b分子结合,可以看作是C 4b/A C 3b的受体。
主要作用:①可竞争性抑制B因子与细胞膜上的C 3b结合,抑制旁路途径C3转化酶(C 3bBbB)在自身细胞膜上形成。
②能从C 4b2b和C 3bBb复合物中快速解离C 2b和Bb,使已形成的C 3转化酶迅速自发衰变,从而阻止膜合物在自身组织细胞膜上形成。
第三节补体受体及其免疫学功能
补体成分激活后产生的裂缝片段,能与免疫细胞表面的特异性受体结合,这对于补体发挥其生物学活性具有重要意义。
补体受体(Complement Receptor,CR)曾按其所结合配体而命名,如C 3b受体,C 3d受体等,但经详细研究后发现,补体受体并非仅与C 3裂解产物反应,还可与补体之外的减分反应,因而又按其发现先后依次命名CR1(CD35),CR2(CD21),CR3(CD11b/CD18),CR4(Gp150/90,CD11c/CD18)。
一、CR1(CD35)
CR1作为免疫粘附(lmmune adgere,ce)受体引起免疫粘附现象。
此受体也称为C3b受体或C3b/C4b受体。
细胞分布:吞噬细胞,B细胞等CR1的免疫功能:
1、中性细胞和单核—巨噬细胞上的CR1,可与结合在细菌或病毒上的C3b结合,促进吞噬细胞的吞噬作用。
2、促进两条激活途径中的C3转化酶(C42,C3bBb)的灭活,这就是补体激活的同时,红细胞不灭溶解的原因,红细胞上有C3b受体C3转化酶没有结合细胞的特异性。
3、作为I因子的辅助因子,促进C3b(和C4b灭活。
4、红细胞上的CR1可与被调理的细菌,病毒或免疫复合物等结合,以便送到肝、脾进行处理。
5、B细胞膜上的CR1与CR2协同作用下,可促使B细胞活化。
二、CR2(CD21)
CR2旧称C3d受体,已证明,它是B细胞上的EB病毒受体。
细胞分布:B细胞等。
功能尚未阐明清楚,但实验表明,当加入CR2配体时可使B细胞活化,据此推想,借结合在Ag—Ab复合物上的C3裂解产物(C3d,C3b),可引起针对该抗原的二次抗体应答。
三、CR3(CD11b/ CD18)
亦称C3b受体,CR3的其它配体还有植物凝集素和细菌脂多糖。
细胞分布:中性粒细胞,单核细胞,吞噬细胞NK细胞。
功能:与吞噬功能有关。
四、CR4(gp150/90,CD 11C/ CD18)
细胞分布为吞噬细胞。
功能:与吞噬功能有关。
第四节补体的生物学活性
补体是在长期的种系进化过程中获得的非特异性免疫因素之一,它也在特异性免疫中发挥作用(见后),它的作用是多方面的。
补体系统的生物学活性,大多是由补体系统激活后产生的裂解产物发挥的。
一、细胞毒及溶菌、杀菌作用
细胞毒作用是指溶细胞的功能,前提是有激活物(主要是Ab)存在的情况下,补体能溶解和杀伤某些G,如霍乱孤菌,杀门氏菌等。
这是经补体的替代途径发挥的作用。
二、调理作用
见前内容C2b C4b
三、免疫粘附作用
免疫复合物激活补体之后,可通过C3b而粘附到表面有C3b受体的白细胞,血小板及某些淋巴细胞上。
形成较大的聚合物,可能有助于被吞噬清除。
四、中和及溶解病毒作用
病毒与相应抗体形成复合物后,加入补体,则明显增强Ab对病毒的中和作用,阻止病毒对宿主细胞的吸附和穿入。
还可溶解有包膜的病毒,据认为是此类病毒包膜上有C1受体。
五、炎症介质作用
炎症也是免疫防御反应的一种表现,感染局部发生炎症时,补体裂解产物可使毛细血管通透性增强吸引白细胞到炎症局部。
(一)激肽样作用
C2a能增加血管通透性,引起失症性充血,具有激肽样作用,故称其为补体激肽。
(二)过敏毒性作用
C3a,C5a
(三)趋化作用
C5a1
复习思考题:
1.试比较补体两条激活途径的主要差异
点.
2.简述补体的生物学功能
3.列出膜结合性补体调节分子的名称,并
简述其作用机制
4.列出体液中可溶性补体调节分子的名称,
并任选其中两种说明其调节作用机制.
5.试述补体系统的组成
6.补体的调理作用和免疫黏附作用是通过
何种成分引起的?其作用机制是什么?
7.补体系统激活后,可产生哪些具有重要
生物学活性的裂解片断,可引发何种生
物学效应?
8.试述补体系统在机体抗感染过程中的作
用.。