克隆基因的表达系统
外源基因原核系统克隆表达的基本流程

外源基因原核系统克隆表达的基本流程
外源基因原核系统克隆表达的基本流程如下:
1. 设计引物:根据外源基因的序列,设计引物,其中至少包括一个启动子和一个终止子。
2. 基因克隆:使用PCR或其他克隆技术,将外源基因与载体DNA连接起来,形成重组质粒。
3. 转化:将重组质粒转化到适当的宿主细胞中,如大肠杆菌。
4. 筛选:通过选择性培养基或其他筛选方法,筛选出带有重组质粒的转化菌落。
5. 培养:将筛选出的转化菌落进行扩增培养,在适当的培养条件下培养细菌。
6. 表达:在培养过程中,外源基因会被宿主细胞转录和翻译,产生目标蛋白质。
7. 提取:收集细菌培养物,利用细胞破裂或其他细胞提取方法,提取目标蛋白质。
8. 纯化:通过各种纯化技术,如柱层析、电泳等,纯化目标蛋白质。
9. 鉴定:利用各种方法,如SDS-PAGE、Western blot等,对
目标蛋白质进行鉴定和定量分析。
10. 应用:利用纯化的目标蛋白质进行后续的研究或应用,如
功能鉴定、结构分析、抗原制备等。
这是一个基本的流程,根据不同的实验目的和具体情况,可能还会涉及到一些其他的步骤和操作。
基因重组技术生产胰岛素

4. 质粒的拷贝数
增加mRNA数量, 提高核糖体与mRNA的结合速度
(1)强启动子提高转录效率;
(2)使用高拷贝表达载体。
可调控的诱导型复制子
宿主大量生长时,抑制载体质粒的复制
宿主大量生长后,诱导载体质粒复制, 增加拷贝数
第31页,共79页。
5. 宿主菌的选择 表达载体与宿主相互匹配
6. 培养条件优化
5’-UAAGGAGG-3’ > 5’-AAGGA-3’
③ SD序列后面的4个碱基
5’-AGGAGGU-3’ ????
如果是A(T),翻译效率最高; 如果是G(C),效率只有50%或25%。
第22页,共79页。
(2)起始密码AUG
AUG左侧的三个碱基也有影响 -半乳糖苷酶的mRNA中:
AUG左侧如果是UAU或CUU时,最为有 效;
(1)培养基
(2)环境因素 (3)诱导时间和剂量
第32页,共79页。
七、原核细胞表达的缺陷
1、缺乏翻译后蛋白质的加工修饰系统, 如糖基化、氨基酸修饰等。
2、原核表达外源蛋白常以包涵体形式存 在,必须经过变性、复性处理才能获 得有生物活性的蛋白,并且其表达量 非常低。
第33页,共79页。
七、原核细胞表达的缺陷
肠杆菌最有效的翻译终止序列。
(3)密码子的选择 ① 受体菌中共表达稀有密码子tRNA基因 ② 将稀有密码子改为偏爱密码子
第28页,共79页。
3. 提高目的产物稳定性 防止被宿主的酶降解。
(1)表达分泌蛋白
外膜 内膜
细胞质
染色体
周间质
质粒
“外排”:蛋白质从细胞质跨过内外膜到培养液中。 “分泌”:蛋白质从细胞质跨过内膜到周间质中。
克隆基因的表达
(2)-35区与-10区之间的距离
这两个保守区间的距离越是接近于17bp,启 动子的活性就越强。
2.翻译起始序列对表达效率的影响 (1)SD序列
SD序列与16SrRNA分子之间的碱基互补程 度,可明显影响mRNA的翻译速度,当序列 为5‘-GGAGG-3’,可与16SrRNA3’端完 全互补,翻译效率最高;而当该序列发生 单碱基突变时,翻译效率会下降30倍。
特定的时间顺序发生,称之为基因表达的 时间特异性。
• 多细胞生物基因表达的时间特异性又称阶 段特异性。
(二)空间特异性 • 在个体生长全过程,某种基因产物在个体
按不同组织空间顺序出现,称之为基因表 达的空间特异性
• 基因表达伴随时间顺序所表现出得这种分 布差异,实际上是由细胞在器官的分布决 定的,所以空间特异性又称细胞或组织特 异性
七、原核表达体系
表达体系的建立包括表达载体的构建、受体细胞的 建立及表达产物的分离、纯化等技术 步骤: 获得目的基因-准备表达载体-将目的基因插入表 达载体中(测序验证)-转化表达宿主菌-诱导 靶蛋白的表达-表达蛋白的分析-扩增、纯化、 进一步检测
(一)获得目的基因
1.对外源目的基因的要求
原核生物缺乏真核生物转录后的加工系统; 同时也缺乏真核生物翻译后的加工系统, 所以目的基因不应具有5’端非编码区以及内 含子结构,只编码成熟的蛋白质或多肽
当需要宿主大量生长时,抑制载体质粒的复制。
当宿主大量生长后,再诱导载体质粒的复制,增 加拷贝数。
6.提高表达产物的稳定性
防止被宿主的酶降解 (1)设计成融合蛋白
克隆基因的表达

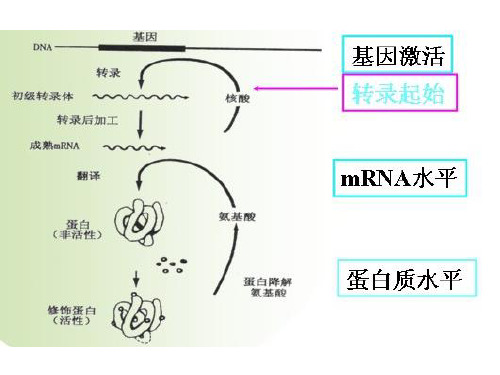
第五章克隆基因的表达基因表达(gene expression)是指储存遗传信息的基因经过一系列步骤表现出其生物功能的整个过程。
典型的基因表达是基因经过转录、翻译,产生有生物活性的蛋白质的过程。
基因的表达主要涉及到两个过程:转录和翻译。
思考:基因表达与克隆基因表达的异同? 从表达的对象、过程、场所等方面考虑。
第一节影响外源基因表达的因素利用基因工程技术高水平表达各种外源蛋白质,无论在理论研究还是实际应用上都是十分重要的。
因此,如何使外源基因在宿主细胞中高效表达,成为基因工程中的关键问题。
应该指出,现在基因工程所涉及的受体细胞多种多样,从细菌到高等动植物细胞,乃至动、植物个体;而所涉及到的基因也是成千上万,各不相同,这种差别使得对特定基因的表达研究就有其特定的个性。
影响外源基因高效表达的各种因素:1. 外源基因的表达效率①启动子的强弱。
有效的转录起始是外源基因能否在宿主细胞中高效表达的关键步骤之一,也可以说,转录起始的速率是基因表达的主要限速步骤。
因此,选择强的可调控启动子及相关的调控序列,是组建一个高效表达载体首先要考虑的问题。
最理想的可调控启动子应该是:在发酵的早期阶段表达载体的启动子被紧紧地阻遏,这样可以避免出现表达载体不稳定,细胞生长缓慢或由于产物表达而引起细胞死亡等问题。
当细胞数目达到一定的密度,通过多种诱导(如温度、药物等)使阻遏物失活,RNA聚合酶快速起始转录。
②核糖体结合位点的有效性。
SD 序列是指原核细胞mRNA 5`端非翻译区同16S rRNA 3` 端的互补序列。
按统计学的原则,一般SD 序列至少含AGGAGG 序列中的4 个碱基。
SD 序列的存在对原核细胞mRNA 翻译起始至关重要。
③SD 序列和起始密码子AUG 的间距。
AUG 是最首选的起始密码子,GUG、UUG、AUU 和AUA有时也用作起始密码子,但非最佳选择。
另外,SD 序列与起始密码子之间的距离以9±3 为适宜。
基因的克隆与表达课件

----TAC -----TTG GAC CTT AAG GAT CCA---
DNA序列
AAT CGG AAG AAT TCA GAC CTA GGT TTA GCC TTC TTA AGT CTG GCT CCA
基因的克隆与表达课件
位相载体----含有3种读码框的系列载体
基因的克隆与表达课件
优点: • 表达效率高 • 产物稳定 • 易鉴定:融合蛋白分子量大,电泳可
➢基因克隆(gene cloning) ➢基因表达(gene expression)
-原核基因表达 -真核基因表达
基因的克隆与表达课件
基因克 隆 Gene Cloning
基因的克隆与表达课件
➢概述 ➢克隆载体 ➢受体细胞 ➢体外重组的策略 ➢基因克隆工作流程
基因的克隆与表达课件
一、概述
• 确定了遗传信息的携带者,即基因的盆 子载体是DNA而不是蛋白质
基因的克隆与表达课件
(二)体外重组 连接体系的建立: • 温度:粘末端连接:12-18℃
平末端连接:室温(低于30℃) • DNA量:载体分子数/目的基因分子数
=1:1-3 • 酶量:平端连接时需加大酶量
基因的克隆与表达课件
(三)转化—Cacl2法、电击法
(四)重组子的筛选及鉴定
1、筛选:平板法(抗生素、蓝白斑)
基因的克隆与表达课件
二.原核生物基因结构和表达特点
基因的克隆与表达课件
• 原核生物染色体DNA是裸露的环形 DNA,其转录和翻译是偶联的连续 进行。
• 原核生物形成多顺反子mRNA: mRNA在合成过程中和多个核糖体 结合,翻译形成多条肽链。
基因的克隆与表达课件
3、一般不含内含子(intron),没有转 录及翻译后加工系统
克隆表达与蛋白质纯化技术

克隆表达与蛋白质纯化技术在生物科学研究领域中,克隆表达与蛋白质纯化技术被广泛应用于蛋白质的生产和研究。
克隆表达是指利用重组DNA技术将目标基因导入宿主细胞,并使其在宿主细胞中表达出来。
蛋白质纯化则是指从克隆表达的细胞中提取和纯化目标蛋白质。
本文将介绍克隆表达与蛋白质纯化技术的基本原理和常用方法。
一、克隆表达技术克隆表达是将感兴趣的基因克隆到表达载体中,通过转染或转化的方式导入细胞中,从而使该基因在细胞内得以表达。
克隆表达技术可分为原核细胞系统和真核细胞系统两类。
1. 原核细胞系统原核细胞系统中,常用的宿主细胞包括大肠杆菌和酵母菌。
在克隆表达中,大肠杆菌是最常用的宿主细胞,其原因在于其繁殖速度快、易于培养和转化、表达效率高等优点。
酵母菌则常用于表达更复杂的蛋白质,因其能够进行糖基化等真核细胞系特有的修饰。
2. 真核细胞系统真核细胞系统中,常用的宿主细胞包括哺乳动物细胞、昆虫细胞和植物细胞等。
哺乳动物细胞系统具有许多优点,如蛋白质修饰和折叠更接近自然情况、大容量表达等,然而其表达成本较高。
昆虫细胞和植物细胞则在表达规模较大的蛋白质时较为常用。
二、蛋白质纯化技术蛋白质纯化是将表达系统中产生的混合蛋白质与其他组分分离的过程,常用的方法有离子交换色谱、亲和层析、凝胶过滤、透析等。
1. 离子交换色谱离子交换色谱是根据蛋白质在离子交换柱中与其反离子交换作用力的不同而进行分离纯化的方法。
常用的离子交换介质有阴离子交换柱和阳离子交换柱。
对于不同电荷性质的蛋白质,可以选择合适的离子交换柱实现分离纯化。
2. 亲和层析亲和层析是利用相互作用力将目标蛋白质与其他组分分离的方法。
常见的亲和层析方法包括金属亲和层析、抗体亲和层析等。
通过对目标蛋白质与特定亲和剂的亲和力进行结合,实现其与其他蛋白质的分离。
3. 凝胶过滤凝胶过滤是利用凝胶材料的大小选择性分离蛋白质的方法。
将混合蛋白溶液经过凝胶柱时,大分子量的蛋白质会被阻滞在柱内,而小分子量的蛋白质则可以通过柱床。
Gateway技术能够克隆一个或多个基因进入到任何蛋白表达系统
Gateway技术能够克隆一个或多个基因进入到任何蛋白表达系统,大大简化基因克隆和亚克隆步骤,同时克隆效率高达95%,当基因在目的表达载体之间快速简便的穿梭时,还可保证正确的方向和阅读框。
Gateway技术基于λ噬菌体位点特异重组系统(attB ×attP →attL ×attR)。
BP和LR两个反应就构成Gateway技术,BP反应利用一个attB DNA片段或表达克隆和一个attP供体载体之间的重组反应,创建一个入门克隆;LR反应是一个attL入门克隆和一个attR目的载体之间的重组反应,用于在平行的反应中把目的序列转移到一个或更多目的载体。
Gateway技术也利用了ccdB选择方法,确保高效率的分离重组克隆。
完成构建Gateway表达克隆仅需两步:⑴创建入门克隆,通过PCR将目的基因克隆进入门载体;⑵混合包含目的基因的入门克隆和合适的目的载体及Gateway LR Clonase,产生表达克隆(用来在合适的宿主中进行蛋白的表达和分析)。
构建入门载体PCR重组克隆重组是从PCR产物创建Gateway入门克隆的一种方法。
通过合并attB位点到上游和下游引物上,然后共同孵育扩增PCR产物和pDONR载体(包含attP位点)及GatewayBP ClonaseTM酶混合物。
接着转化进大肠杆菌中,您将会获得包含目的基因的入门克隆,同时目的基因两侧具有attL重组位点。
这个入门克隆可以与任何Gateway目的载体进行重组。
表达系统一旦您构建好Gateway入门克隆,就可以使用Gateway技术进入到几乎是无数种的表达系统。
Gateway技术为进行基因功能分析和蛋白表达提供了广泛深入的方法,通过把目的基因转移到优化构建的体外表达载体,pEXP1-DEST 或pEXP2-DEST,然后开始体外蛋白合成。
目前有很多具有不同启动子的大肠杆菌目的载体,提供一系列表达选择,Gateway目的载体也提供N端或/和C端融合标签的选择,简化表达蛋白的纯化和检测。
简述外源基因原核系统克隆表达的基本流程
简述外源基因原核系统克隆表达的基本流程外源基因在原核系统中的克隆表达是通过一系列步骤来实现的。
以下是基本的流程:1. 选择质粒载体(Plasmid Vector):-选择一个合适的质粒,通常是圆形DNA 分子,具有自主复制的能力。
质粒通常包含选择标记(例如抗生素抗性基因)和表达调控元件(例如启动子、终止子等)。
2. 准备目标基因:-获取外源基因,这可以是从其他生物中克隆得到的DNA 片段。
这个基因应该编码所需的蛋白质或RNA。
3. 限制性内切酶切割:-使用限制性内切酶切割质粒载体和目标基因。
选择适当的酶,以确保两者切口相互兼容。
4. 连接(Ligation):-将切割后的质粒和目标基因连接在一起,形成重组质粒。
这一步通常涉及DNA 连接酶。
5. 转化(Transformation):-将重组质粒导入宿主细菌中。
这可以通过热激冲击、电穿孔或其他方法实现。
质粒包含抗生素抗性基因,使得只有带有重组质粒的细菌能够在含有抗生素的培养基中生长。
6. 筛选(Screening):-鉴定带有正确重组质粒的细菌。
这可以通过PCR、酶切鉴定等技术来进行。
7. 培养:-将筛选出的正常克隆株培养起来,以增大其数量。
8. 表达:-利用宿主细菌的生物机制,使得外源基因在细菌中表达。
这通常涉及到适当的启动子和终止子,以及其他调控元件。
9. 纯化:-如有必要,对表达的蛋白质进行纯化。
这可以通过各种方法,如层析、电泳等来实现。
整个流程的成功依赖于实验室技术的熟练操作和对基因工程原理的深刻理解。
这些步骤的每一步都需要谨慎操作,以确保最终得到具有期望表达产物的克隆。
基因工程的表达系统
基因工程的表达系统
基因工程是一门研究在实验室中人为操纵和修改生物体遗传材料的学科,从而达到改变或改造生物性能的目的。
其中,表达系统是基因工程的重要技术之一,也是用来实现基因功能分析和基因转录的重要手段。
表达系统是指将外源基因引入宿主菌株,进而在宿主菌株中实现外源基因表达的技术系统。
表达系统包括各种实验技术,如基因克隆、基因表达定位、基因调控、基因表达调控、基因表达产物分离等技术。
基因克隆是在表达系统中要完成的第一步,即将指定的DNA序列导入宿主菌株,这一步可以使用质粒克隆技术或集成克隆技术来实现,这两种技术都简单、快捷、可靠,因此在基因工程的实验中得到了广泛的应用。
定位和调控是表达系统中的第二步,目的是将克隆好的外源基因放置在宿主菌株中能够正常发育和表达的位置,以便获得正确的表达方式,这一步可以使用启动子技术、启动子组装技术、表达调控因子技术等来实现。
表达系统的最后一步是表达产物的分离,也就是将克隆好的外源基因在宿主菌株中表达出来的产物进行分离,这一步可以使用浓缩、沉淀、超滤、离心分离等方法来实现,以获得更高的产物纯度。
总的来说,基因工程的表达系统是一整套实验技术,既可以用于表达和功能分析基因,也可以用于产生新型药物、新型酶、新型农药、新型食品添加剂等多种产品,是基因工程的重要技术手段。
简述基因工程中常用的表达系统及其优缺点
简述基因工程中常用的表达系统及其优缺点基因工程是现代生命科学的主要分支之一,这门学科的发展主要是基于分子生物学的基础知识,致力于研究操纵和编辑生物体的遗传物质,以实现一定的目的。
基因工程在农业、医疗、军事、安全和环境保护等诸多领域的应用,已经给人类带来了许多好处。
基因工程中使用的表达系统主要有质粒、细菌表达系统、动物细胞表达系统和植物细胞表达系统。
它们各自具有独特的优缺点,可以满足不同的基因工程应用。
一、质粒表达系统质粒表达系统是基因工程中最常用的表达系统之一,它可以在宿主细胞中对外源基因进行稳定表达。
传统的质粒表达系统由一个含有增强子的外源基因组成,这些增强子可以有效地增强基因的表达,使基因的表达水平有所提高。
优点是这种表达系统的准备工作容易、成本低,表达效率较高,但是缺点也很明显,宿主细胞往往会产生抗性,导致基因表达受到抑制,这也是这种表达系统的主要缺点。
二、细菌表达系统细菌表达系统是一种非常常用的表达系统,主要是利用细菌的表达机制来表达外源基因。
优点在于它可以有效地表达基因,并且表达的成本也比较低,而且它的可制备性也很高,在细菌中表达的成本也比较低,但是也有缺点,如细菌中表达的基因通常缺乏活性,使得基因不能有效地表达。
三、动物细胞表达系统动物细胞表达系统是基因工程中最常用的表达系统之一,主要是利用动物细胞的代谢机制,将外源基因植入动物细胞中,再使其表达出目标基因t。
这种表达系统的优点有:(1)物细胞可以制备细胞因子,从而使表达的基因更具有活性;(2)动物细胞具有复杂的细胞调控机制,可以有效地表达基因;(3)动物细胞可以比较容易地进行克隆,这也是这种表达系统最有价值的地方。
但是,动物细胞表达系统也存在一些缺点,比如宿主细胞抗性增加,基因表达受到抑制;高细胞表达成本,细胞的获得和细胞的维护成本都很高;动物细胞的移植技术较弱,缺乏稳定的移植技术。
四、植物细胞表达系统植物细胞表达系统是基因工程中最近发展起来的表达系统,主要是利用植物细胞的代谢机制来表达外源基因。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
(3) SD序列与起始密码子之间的距离的影响:
SD序列与起始密码子之间的距离是影响mRNA转译成 蛋白的主要因素之一。在很多情况下,SD序列位于 AUG之前大约七个碱基处。在此间隔中少一个碱基 或多一个碱基,均会导致翻译起始效率不同程度的降 低
密码子偏爱的影响 翻译终止的影响
终止密码子对翻译的效率有很大影响. UAA,UAG,UGA
4 表达系统对表达产物的影响
表达载体和受体细胞共同组成了外源基因 的表达系统.
二、外源基因在原核细胞中的表达
Operator promoter
Structural gene
terminator
G1 G2
(2) SD序列与起始密码子之间的序列的影响
SD序列下游的碱基若为AAAA或UUUU,翻译 效率最高;而CCCC或GGGG的翻译效率则分别 是最高值的50%和25%。紧邻AUG的前三个 碱基成份对翻译起始也有影响,对于大肠杆菌b半乳糖苷酶的mRNA而言,在这个位置上最佳 的碱基组合是UAU或CUU,如果用UUC、 UCA或AGG取代之,则酶的表达水平低20倍
2.Shine-Dalgarno(SD)序列
位于翻译起始密码子(AUG)上游5-13bp处 的由3-9个bp组成的核苷酸序列富含嘌呤 核苷酸, 5’ UAAGGAGG 3’,,它通 过识别大肠杆菌核糖体小亚基中的16S rRNA 3’端区域3’ AUUCCUCC 5’并与 之专一性结合,将mRNA定位于核糖体 上,从而启动翻译;
启动子 :是DNA上的能与RNA聚合酶结合并能起始 mRNA合成的序列 ,是基因表达不可缺少的序列.
保守序列
-10区(Pribnow框)也称TATA-box,TATAAT
-35区(Sextama框)TTGACA,是RNA聚合酶σ 亚基的识别与结合位点
-1 +1
+30
TTGACA
15~20bp TATAAT
6bp
CAT
- 35元件
- 10元件
启动子
启动子
-35 区序列
PlL PrecA PtraA Ptrp Plac Ptac
TTGACA TTGATA TAGACA TTGACA TTTACA TTGACA
Ptac = 3 Ptrp = 11 Plac
-10 区序列
GATACT TATAAT TAATGT TTAACT TATAAT TATAAT
衰减子
是原核生物基因表达调控的精细调节装置, 是基因表达的负调控元件.
3 翻译过程对表达的影响
翻译是mRNA指导多态链合成的过程,包括 多态链合成的起始,延伸和终止过程
翻译起始对基因表达的影响
原核生物起始密码子(AUG),SD,SD序列与起始密码之间的 距离保持在9±3个核苷酸;真核生物的5’端帽子结构,起始密 码子周边共有序列,有效的ORF是翻译所必需
双向作用 提高转录效率 无位置效应 远距离作用 组织特异性:只能在特定组织中增强与其相连的
启动子的活性 无基因特异性
沉默子
远距离调节启动子以降低转录效率的一段 DNA序列,是调节基因表达的顺势作用负 调控元件,最早在酿酒酵母中发现
终止子 Terminator
为转录提供终止信号的一段DNA序列,是 基因表达的顺势作用负调控元件.
克隆基因的表达系统
基因的表达:指遗传信息从DNA到RNA再到 蛋白质的过程
Operator promoter
Structural gene
terminator
G1 G2
G3
一、影响外源基因表达的因素
1、阅读框架:指基因的编码区,包含从起始密码 子至翻译终止密码子之间的cDNA序列。
2、顺式作用元件 顺式作用元件对基因转录起始的调控 ➢ 启动子 ➢ 增强子 ➢ 沉默子 顺式作用元件对转录终止的调控 ➢ 终止子 ➢ 衰减子
外源基因与表达载体连接后形成正确的开放 阅读框架(open reading frame)。
必须利用原核细胞的调控原件(启动子等) 防止外源基因产物对宿主细胞的毒害
(二)、原核生物基因表达的调控序列
原核生物基因表达的调控序列主要涉及启 动子,S-D序列,终止子,衰减子等序列
1.原核生物启动子结构
G3
(一)、原核生物基因表达的特点
➢ 只有一种 RNA 聚合酶,识别原核细胞的启动子,催化所 有的RNA合成。
➢ 以操纵子为单位,数个相关的结构基因与其调控区结合 形成一个表达的协同单位
➢ 转录和翻译偶联、连续进行。 ➢ 不含内含子,缺乏转录后的加工系统 ➢ 调控主要在转录水平上 ➢ mRNA的核糖体结合位点上含有一个转译起始密码子及
启动子
是RNA聚合酶识别,结合并起始转录的一段 DNA序列,长度随生物种类不同而异.主要 特征:
序列特异性: 方向性:结构一般是不对称的,正反两个
方向只有一种具有启动功能 位置特异性 种属特异性:
增强子
远距离调节启动子并提高转录效率的一段DNA序列, 由多个元件组成,每个元件可与一种或多种转录调 控因子结合.最早在SV40中发现,作用方式特点:
核糖体结合位点对外源基因表达的影响
(1) SD序列的影响:
一般来说,mRNA与核糖体的结合程度越强,翻译的起 始效率就越高,而这种结合程度主要取决于SD序列与 16S rRNA的碱基互补性,其中以GGAG四个碱基序 列尤为重要。对多数基因而言,上述四个碱基中任何 一个换成C或T,均会导致翻译效率大幅度降低
乳糖启动子lac---来自大肠杆菌的乳糖操纵子 色氨酸启动子trp-来自大肠杆菌的色氨酸操纵子 PL和PR启动子 -l噬菌体中得到的一类启动子 比lac启动子的活性高8-10倍,比trp启动子活性高.
-10 区顺序:GATAAT, -35区顺序:TGACTA
tac启动子
由trp 和 lac启动子人工构建的杂合启动 子
同16S核糖体RNA末端互补的序列,即Shine-Dalgarno (S-D)sequence
mRNA上一段同核糖体16S RNA3’末端碱基互补的序列,叫 Shine-Dalgarno(S-D)序列
原核表达系统的注意事项
通过表达载体将外源基因导入宿主菌,并指导 宿必须用cDNA或全 化学合成基因