质粒载体构建
实验四.重组质粒载体的构建

5'…G↓AATTC…3' →5'… G AATTC…3' 3'…CTTAA↑G …5' →3'… CTTAA G…5'
DNA纯度、缓冲液、温度条件及限 制性内切酶本身都会影响限制性内切酶 的活性。大部分限制性内切酶不受RNA 或单链DNA的影响。当微量的污染物进 入限制性内切酶贮存液中时,会影响其进 一步使用,因此在吸取限制性内切酶时,每 次都要用新的吸管头。如果采用两种限 制性内切酶,必须要注意分别提供各自的 最适盐浓度。若两者可用同一缓冲液,则 可同时水解。若需要不同的盐浓度,则低 盐浓度的限制性内切酶必须首先使用,随 后调节盐浓度,再用高盐浓度的限制性内 切酶水解。
2、 混匀反应体系后,将eppendorf管置于适 当的支持物上(如插在泡沫塑料板 上),37℃水浴保温2-3小时,使酶切反应 完全。 3、 每管加入2μl 0.1mol/L EDTA(pH8.0), 混匀,以停止反应,置于冰箱中保存备用。
注意事项
1. 酶切时所加的DNA溶液体积不能太大,否则DNA溶液中其他成 分会干扰酶反应。 2、酶活力通常用酶单位(U)表示,酶单位的定义是:在最适反应 条件下,1小时完全降解1mg lDNA的酶量为一个单位,但是许多 实验制备的DNA不象lDNA那样易于降解,需适当增加酶的使用 量。反应液中加入过量的酶是不合适的,除考虑成本外,酶液中 的微量杂质可能干扰随后的反应。 3、市场销售的酶一般浓度很大,为节约起见,使用时可事先用酶 反应缓冲液(1×)进行稀释。另外,酶通常保存在50%的甘油 中,实验中,应将反应液中甘油浓度控制在1/10之下,否则,酶 活性将受影响。
构建DNA限制性内切酶图谱有许多 方法。通常结合使用多种限制性内切酶, 通过综合分析多种酶单切及不同组合的 多种酶同时切所得到的限制性片段大小 来确定各种酶的酶切位点及其相对位置。 酶切图谱的使用价值依赖于它的准确性 和精确程度。
质粒载体的操作和cDNA文库的构建

(一)细菌培养物的生长从琼脂平板上挑取一个单菌落,接种到培养物中(有含有行当抗生素的液体培养基中生长),然后从中纯化质粒,质粒的提纯几乎总是如此。
现在使用的许多质粒载体(如pUC系列)都能复制到很高的拷贝数,惟致只要将培养物放在标准LB 培养基中生长到对数晚期,就可以大量提纯质粒。
此时,不必造反性地扩增质粒DNA。
然而,较长一代的载体(如pBR322)由于不能如此自由地复制,所以需要在得到部分生长的细菌培养物中加入氯霉素继续培养若干小时,以便对质粒进行性扩增。
氯霉素可抑制宿主的蛋白质合成,结果阻止了细菌染色体的复制,然而,松弛型质粒仍可继续复制,在若干小时内,其拷贝数持续递增。
这样,像pBR322-类的质粒,从经氯霉素处理和未经处理的培养物中提取质粒的产量迥然不同,前者大为增高。
多年来,加入足以完全抑制蛋白质合成的氯霉素μg/ml)已成为标准的操作、用该方法提取的质粒DNA量,对于分子克隆中几乎所有想象到的工作任务。
(二)细菌的收获和裂解细菌的收获可通过离心来进行,而细菌的裂解则可以采用多种方法中的任意一种,这些方法包括用非离子型或离子型去污剂、有机溶剂或碱进行处理及用加热处理等。
选择哪一种方法取决于3个因素:质粒的大小、小肠杆菌菌株及裂解后用于纯化质粒DNA的技术。
尽管针对质粒和宿主的每一种组合分别提出精确的裂解条件不切实际,但仍可据下述一般准则来选择适当方法,以取得满意的结果。
1)大质粒(大于15kb)容易受损,故应采用漫和裂解法从细胞中释放出来。
将细菌悬于蔗糖等渗溶液中,然后用溶菌酶和EDTA进生处理,破坏细胞壁和细胞外膜,再加入SDS一类去污剂溶解球形体。
这种方法最大限度地减小了从具有正压的细菌内部把质粒释放出来所需要的作用力。
2)可用更剧烈的方法来分离小质粒。
在加入EDTA后,有时还在加入溶菌酶后让细菌暴露于去污剂,通过煮沸或碱处理使之裂解。
这些处理可破坏碱基配对,故可使宿主的线状染色体DNA变性,但闭环质粒DNA链由于处于拓扑缠绕状态而不能彼此分开。
质粒的构建

② -半乳糖苷酶Xgal显色反应: -半乳糖苷酶能把无色的化合物 Xgal分解成半乳糖和一个深蓝色的 物质5-溴-4-氯靛蓝。 -半乳糖苷酶 Xgal 半乳糖
5-溴-4-氯靛蓝
③ lacZ的肽互补 1)-肽( lacZ’ ): -半乳糖苷酶N端的一段氨基酸片断 (11-41氨基酸)。 N端的11-41aa N端的11-41aa N端的11-41aa N端的11-41aa C端大部分 C端大部分 C端大部分 C端大部分
(2)长度
4363bp
(3)选择标记 氨苄青霉素和四环素抗性。
(4)克隆位点
24个克隆位点。
其中9个会导致Tetr基因失活(如 BamH I、Hind Ⅲ、Sal I); 3个会导致Ampr基因失活(Sca I、 PvuI、Pst I)。
(5)pBR322的筛选 ①双抗菌素抗性选择标记
在EcoRⅠ和HindⅠ酶剪切位点之间插入外源基因
(2)长度
(3)克隆位点
约2.7kb
10个连续的单一限制酶切位 点,位于lacZ’基因的5’端。
ห้องสมุดไป่ตู้
pUC18/19
选择标记 Ampicillin 抗性和 lacZ的肽互补(蓝白 斑)相结合。 筛选方法
选用带有含有ampicillin和X-gal的培养基 受体菌lacZ突变(lacZ∆M15)
受体菌lacZ突变(lacZ∆M15) 受体菌基因组的-半乳糖苷酶基因的 氨基端有缺失(缺失肽),不能形成 4聚体的活性酶,不能分解Xgal 受体菌株:JM系列、TG1、TG2、 XL1-blue、XS127、XS101、KK2186、 MV1184、DH5a
质粒载体生物学特征
分类 按复制类型可分为:松弛型 严紧性 按转移性可分为转移型和非转移型
重组表达质粒的构建——原核表达载体选择
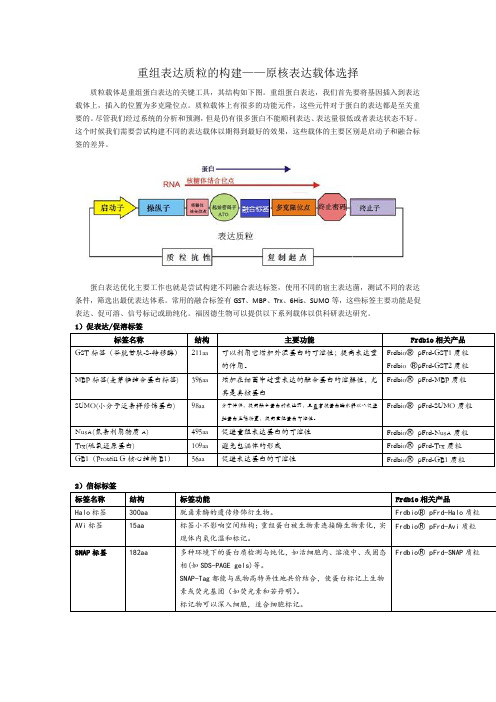
重组表达质粒的构建——原核表达载体选择质粒载体是重组蛋白表达的关键工具,其结构如下图。
重组蛋白表达,我们首先要将基因插入到表达载体上,插入的位置为多克隆位点。
质粒载体上有很多的功能元件,这些元件对于蛋白的表达都是至关重要的。
尽管我们经过系统的分析和预测,但是仍有很多蛋白不能顺利表达、表达量很低或者表达状态不好。
这个时候我们需要尝试构建不同的表达载体以期得到最好的效果,这些载体的主要区别是启动子和融合标签的差异。
蛋白表达优化主要工作也就是尝试构建不同融合表达标签,使用不同的宿主表达菌,测试不同的表达条件,筛选出最优表达体系。
常用的融合标签有GST、MBP、Trx、6His、SUMO等,这些标签主要功能是促表达、促可溶、信号标记或助纯化。
福因德生物可以提供以下系列载体以供科研表达研究。
1)促表达/促溶标签2)信标标签3)纯化标签我们选择表达载体的时候不但要考虑蛋白怎么表达成功,更要考虑蛋白怎么纯化出来,纯化的问题主要是考虑纯化标签和酶切位点的选择,下表我们列举了常见的纯化标签和酶切位点。
4)酶切位点以上为原核表达常用的标签和酶切位点,其性质也都作了简要的介绍,各专业网站或专业书籍已对此做详尽解释,科研工作者可根据具体实验设计方案,组合设计以上标签和酶切位点的使用。
特别值得注意的是,选用和设计蛋白酶切位点的时候首要考虑的是序列内部有没有蛋白酶位点,同时要考虑酶切的效率和蛋白酶试剂成本。
一般商业化载体,在标签蛋白与载体多克隆位点之间都设计有酶切位点。
标签可设计在N-端也可在C-端,设计在N-端的优势是,可通过标签高效翻译起始位点带动插入蛋白的表达,可溶性标签的高效表达更可促进蛋白的可溶性表达;同时,大部分的蛋白内切酶的切割位点在C-端,所以标签设计在N-端可将标签切割完全。
在设计标签序列与酶切位点的时候还要考虑N-端稳定性原则,也就是所谓宿主细胞的N-端规则(N-end rule),这个要避免;同时,还应该检查是否引入了可与别的蛋白相互作用的序列或者蛋白酶切位点。
载体构建介绍

5.常见问题
选择标记类型和选择 1、选择性标记类型 药物抗性(如Kan,Amp等) 营养依赖性标记(如SC-Leu 等) 2、如何选择 根据所选载体上所带的标记
5.常见问题
Gateway系统引物设计-需要对读码框
这是因为中间载体和终载体上某些编码氨基酸或者抗性 基因同目的片段共用一个起始密码子。
载体构建
戎浩 2015.12
C
ONTENTS
目录
1 2 3 4
载体简介 载体构建
注意事项 常见问题
1.载体简介
目的基因的克隆与鉴定
ห้องสมุดไป่ตู้
生理检测 纯化
载体构建
大肠转化,质粒 提取与鉴定
移栽
分子检测
继代繁殖 农杆菌的转化与活化
外植体制备
筛选
浸
染
共培养
1.1载体
载体(vector) ,能将外源DNA或基因片段携带入 宿主细胞内的一个具有自我复制能力的DNA分子。
退火温度不合适,设置梯度,选择最适退火温度
2、条带不单一
引物不特异,适当增长引物序列长度;
适当提高退火温度
5.常见问题
载体酶切的问题 1、酶切质粒浓度和纯度要好 2、酶切温度和时间
如果两个酶的最适温度不同,建议单酶切,回收后在
用另一个酶切,时间最好过夜切。 3、没有切开 可能是酶失活,建议酶切时增加阳性对照,确定酶是 否好用
5.常见问题
克隆基因的酶切位点及引物问题 1、单酶切 单酶切后进行连接,质粒自连、目的片段自连、目的
片段之间连接、目的片段和载体各种错误连接、目的片段
反向连接等等,尽量不选单酶切 2、保护碱基数目的问题。 在设计PCR引物时,引入酶切位点后,常常要加入保 护碱基,这会使得酶切效率大大提高。
质粒载体的构建

四、连接 很多公司都有快速连接试剂盒和普通试剂盒。一般都是 16 度数个小时或者过夜,或者 4 度 过夜,不一而同。曾在园子里看到一个高人的文章,认为是时间越长越好,甚至建议 4 度放 几天,我没有试过。16 度可以用 PCR 仪创造,或者找一个泡沫盒,加上冰,水搞到 15 度 左右,放入 4 度冰箱即可。我用的 TAKARA 的快速连接试剂盒,4 度过夜。 菜鸟体贴提示: 1、 通过电泳,粗略判定质粒和 DNA 的浓度比例。一般加入连接液的 DNA:质粒=9:1 2、 放入质粒和 DNA 的总量要适当,不可超过说明书,宁少不要多。
质粒载体的构建-菜鸟入门手册
dongkey 制作
本人刚刚完成质粒载体的构建,总结了一下,以方便要做这方面的菜鸟同学借鉴一下,希望 老鸟同学批评指教。 一、确定插入的基因片段。 首先要确定自己要插入载体中的基因片段,比如要做蛋白表达和功能,可以选择 CDS 区进 行插入。然后自然是如何得到这段基因片段的问题了。最常见的是 PCR 解决了。那么就涉 及到引物的设计。插入 CDS 基因片段的例子:找到 CDS,根据 CDS 设计全长引物,然后 加上内切酶的碱基片段(这要根据自己手头上有的质粒的内切酶位点来决定了),注意,前 面要加上保护碱基!然后进行 PCR。 PCR 后要跑电泳确定目的基因的长度是正确的。 用乙醇沉淀法纯化 PCR 产物(具体见分子克隆一书) 菜鸟体贴提示:要在质粒上找到两个最好不是连续在一起的内切酶,然后分别添加在两条引 物的 5‘端,当然内切酶的温度最好是一致的,而且可以能够同时切开的(双酶切)。这可 以在试剂公司的限制性内切酶的列表上找到有没有共同的 BUFFER 及其双酶切时的活性如 何(当然是越大越好了)。内切酶的公司一般可以找 NEB, TAKARA, TOYOBO 等公司,本 人用的是 TAKARA。 二、酶切 将质粒和基因片段分别进行酶切。最省事的是同时双酶切了。一般酶切温度都是 37 度。 菜鸟体贴提示: 1、 确定质粒和基因片段的量!!(宁少不多),根据说明书加样,一般是 DNA+BUFFER+
载体构建介绍
1.2载体的功能
运送外源基因高效的转入到受体细胞中 为外源基因提供复制能力或整合能力 为外源基因的扩增或表达提供条件
1.3载体应具备的条件
具有对受体细胞的可转移性 具有与特定受体细胞相适应的复制位点或整合位点 具有多种单一的酶切位点 具有合适的选择性标记
2.载体构建(酶切连接)
3.3 PCR
为获得和目的基因完全相同的序列,一般使用高保真酶 进行目的片段的扩增。(加尾)
3.4 PCR产物纯化
目的片段
琼脂糖凝胶电泳
切胶回收(方法参照试剂盒)
1.避免一些非特性条带 2.防止PCR体系中一些成分对转化的影响
3.5 BP反应
3.6 大肠杆菌转化 3.7 质粒提取
3.8 LR反应
现在人们还在不断寻找新的载体,如叶绿体或线粒体 DNA也有可能成为载体。
1.2载体的分类
克隆载体
植 物 基 中间载体 因 工 程 卸甲载体 载 体
表达载体ቤተ መጻሕፍቲ ባይዱ
一元表达载体 双元表达载体
1.2载体分类
克隆载体
可携带出入的外源 DNA片段并可转入受体 细胞中大量扩增的DNA 分子。通常含有筛选 标记,在插入外源的 基因的时候不会破坏 载体本身的自我复制 功能。
1.2载体分类
中间载体
1.2载体分类
卸甲载体
野生型的Ti质粒不能直接作为基因工程的载体,期TDNA区存在一段阻碍细胞分化和植株再生的基因Onc, 将其切除,即“解除”其“武装”,因此称为卸甲载 体。
1.2载体分类
双元表达载体
一般植物表达载体 是成套使用的,一套 中有两个,一个是带 有可以供插入外源表 达基因的MCS和筛选 标签的融合蛋白(通 常是GFP,GUS或抗性 基因蛋白)的普通表 达载体。另一个载体 就是双元表达载体。
质粒载体的构建及其在基因工程中的应用研究
质粒载体的构建及其在基因工程中的应用研究随着基因工程技术的不断发展,质粒载体成为了实现基因转化的重要手段之一。
质粒载体构建是基因工程研究的重要环节,其成功与否直接关系到后续研究的开展和成果的取得。
本文将介绍质粒载体的构建原理、构建方法以及其在基因工程中的应用研究。
一、质粒载体的构建原理质粒载体是人工合成的单链圆形DNA分子,作为存储DNA序列的一种手段,其基本结构特征主要包括起始端、终止端、多个酶切位点、标记位点等。
考虑到质粒载体在基因工程研究中的应用,构建原理主要包括以下几点:1. 合成目的基因的DNA序列,包括启动子、编码区、终止子等。
2. 找到合适的质粒载体,通过酶切识别并将目的基因DNA序列与质粒载体进行连接。
3. 对连接后的质粒载体进行转化,将其转移到细胞中,获得外显表达的蛋白。
质粒载体的构建原理相对简单,但质粒载体在实际应用中的构建过程则需要复杂的技术手段和严密的实验操作。
二、质粒载体构建方法1. 基于PCR扩增法PCR扩增法是目前基因工程研究中最常用的方法之一。
选择需要进行扩增的目的基因DNA序列,使用酶切产生目的碎片,通过PCR扩增后,利用限制性内切酶等酶切方法进行连接。
2. 基于化学合成法在基于化学合成法中,研究者可以通过化学合成方式来合成目的基因DNA序列,在此基础上通过PCR扩增和限制性内切酶等酶切方法进行连接。
3. 基于网站选择法基于网站选择法是现在比较流行的方法之一,具有操作简单、成本低等优点。
研究者可以在网站上选择目的基因序列,并结合实验室已有的质粒载体库,在网站上进行设计、合成、定制和质粒表达等步骤。
三、质粒载体在基因工程中的应用研究质粒载体在基因工程中的应用研究十分广泛,可以用于植物基因转化、动物基因转化、疫苗研发、DNA疫苗的制备、表达蛋白的研究等方面。
1. 植物基因转化通过基因转化技术向植物中加入外源基因,可以使植物表现出新的性状或特征。
在实践中,研究者会将需要转入的序列合成后,利用限制性内切酶或其他酶切方法将其插入到质粒载体中,再利用农杆菌等工具将质粒载体导入植物细胞中,从而实现植物基因转化。
载体构建质粒构建步骤有哪些?
载体构建质粒构建步骤有哪些?载体构建过程:1、引物设计2、⽬的⽚段选取:RNA提取、RNA反转录、PCR扩增、PCR产物纯化3、双酶切4、连接:T4 DNA ligase连接或者同源重组连接(新贝⽣物:#B101、#B102)5、转化6、菌落PCR7、测序:1)摇菌;2)送样; 3)⽐对;8、菌种保存:菌种⽐对成功,则可保存菌种备⽤。
9、质粒提取:菌种⽐对成功,冻存菌种后,菌液⽤于提取质粒。
⼀、载体构建基本原理分、切、连、转、筛1、分:分离出要克隆的⽬的基因及载体。
2、切:⽤限制性内切酶切割⽬的基因和载体,使其产⽣便于连接的末端。
限制性内切酶:是⼀类能识别双链DNA中特定碱基顺序的核酸⽔解酶。
限制性核酸内切酶根据识别切割特性,催化条件及是否具有修饰酶活性分为三⼤类。
其中Ⅱ型酶能识别双链DNA的特异顺序,并在这个顺序内切割,产⽣特异性DNA⽚段,是DNA 重组技术中常⽤的酶。
Ⅰ型酶:具有修饰和切割功能,⽆固定切割位点Ⅲ型酶与Ⅰ型类似,能识别特异位点,但切割位点在识别位点以外Ⅱ型酶特点:①识别顺序⼀般为4-6个碱基对②识别顺序具有180度的旋转对称性,呈完全的回⽂结构③Ⅱ型酶对双链DNA两条链同时切割,可产⽣两种不同末端:平末端,粘末端平末端:在识别顺序的对称轴上,对DNA同时切割形成平末端,如:SmaI5’-CCC GGG-3’ 5’-CCC GGG-3’3’-GGG CCC-5’ 3’-GGG CCC-5’5′突出粘末端:在识别序列的两侧末端切割DNA双链,于对称轴的5 ′末端切割产⽣5 ′端突出的粘性末端,如:Hind Ⅲ5’―AAGCTT―3’ 5’― A 5’-AGCTT―3’3’―TTCGAA―5’ 3’― TTCGA-5’ A―5’3′突出粘末端:与5′突出粘末端作⽤相反,产⽣3 ′端突出粘末端,如:PstI5’―CTGCAG―3’ 5’―CTGCA-3’ G―3’3’―GACGTC―5’ 3’―G 3’-ACGTC―5’3、连:将切割后的⽬的基因和载体⽤T4 DNA连接酶连接或者同源重组⽅法连接。
载体构建简介
载体构建SOP载体构建(vectorconstruction),为把DNA分子运送到受体细胞中去,必须寻找一种能进入细胞、在装载了外来的DNA片段后仍能照样复制的运载体。
理想的运载体是质粒(plasmid),在基因工程中,常用人工构建的质粒作为载体。
载体构建即是构建含外源DNA的质粒。
载体构建是分子生物学研究常用的手段之一。
主要包括已有载体多克隆位点MCS的改造和已有载体启动子、增强子、筛选标记等功能元件的改造。
载体构建完成后可利用PCR原理进行测序验证。
原核重组表达常用载体构建策略有多种选择,(1)常用的酶切连接是较为广泛使用的克隆技术,主要优点是技术稳定,缺点是周期长、步骤多,任何一个环节产生的误差都会影响克隆构建的失败,如用酶切连接的策略进行载体批量构建,不同载体和不同外源基因尽可能选用相同的上下游酶切位点,需特别留意的是基因内部不能有与上述相同的酶切位点;(2)同源重组是目前流行的克隆技术。
同源重组(Homologous Recombination)是指发生在非姐妹染色单体(sister chromatid)之间或同一染色体上含有同源序列的DNA分子之间或分子之内的重新组合。
该技术可以任意载体任意基因片段快速实现多片段长片段定点定向克隆,最大的优点在于不依赖于酶切位点进行克隆,操作时间也非常短,将目的片段和线性载体按照一定比例混合后,在重组酶的作用下即可发生重组,本实验室常用37℃的温度,30min反应即可完成。
一、目的载体进行双酶切(一)实验准备1.实验材料:质检合格的目的载体。
2.试剂与耗材:buffer,ddH2O,琼脂糖(Spain,9012-36-6)、TBE缓冲液、PCR 管(国产,81245)、1.5ml离心管(国产)、枪头(国产)、Axygen AxyPrep PCR cleanup Kit(Axygen,AP-PCR-250)、限制性内切酶A、限制性内切酶B。
3.仪器与设备:PCR仪(杭州博日,TC-96/G/H(b)A)、mini离心机(其林贝尔,LX600)、电泳槽(北京君意,JY-SPCT)、凝胶成像仪(Bio-rad,GelDoc/ChemiDocuniversal hood II)、分光光度计(天根,OSE-260-01)。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
附录A 表达载体的构建
1、大肠杆菌质粒DNA的提取(SDS碱裂解法)
该方法参照分子克隆指南的方法修订而成。
细胞的制备
(1) 将2ml含相应抗生素的LB培养基加入到容量为15ml并通气良好(不盖紧)的试管中,然后接入单菌落,于37℃剧烈振摇下培养过夜。
(2) 将1.5ml培养物倒入微量离心管中,用微量离心机于4℃以最大转速离心30s,将剩余的培养物贮存于4℃。
(3) 吸去培养液,使细菌沉淀尽可能干燥。
细胞的裂解
(4) 将细菌沉淀重悬于100μl用冰预冷的碱裂解液I中,剧烈振荡。
(5) 加200μl新配制的碱裂解液Ⅱ于每管细菌悬液中,盖紧管口,快速颠倒离心管5次,以混合内容物。
应确保离心管的整个内表面均与碱裂解液Ⅱ接触。
不要振荡,将离心管放置于冰上。
(6) 加150μl用冰预冷的碱裂解液Ⅲ,盖紧管口,反复颠倒数次,使溶液Ⅲ在粘稠的细菌裂解物中分散均匀,之后将管置于冰上3-5min。
(7) 用微量离心机于4℃以最大转速离心5min,将上清转移到另一离心管中。
(8) 加等量体积的氯仿:异戊醇(24:1),振荡混合有机相和水相,然后用微量离心机于4℃以最大转速离心5min,将上清转移到另一离心管中。
质粒DNA的回收
(9) 用2倍体积的乙醇于室温沉淀核酸。
振荡混合,于室温放置2min。
(10) 用微量离心机于4℃以最大转速离心5min,收集沉淀的核酸。
(11) 小心吸去上清液,将离心管倒置于一张纸巾上,以使所有液体流出,再将附于管壁的液滴除去。
(12) 加1ml 70%乙醇于沉淀中并将盖紧的离心管颠倒数次,用微量离心机于4℃以最大转速离心2min,回收DNA。
(13) 小心吸去上清液,将离心管倒置于一张纸巾上,以使所有液体流出,再将附于管壁的液滴除去。
(14) 将开口的离心管置于室温使酒精挥发,直至离心管内没有可见的液体存在(5-10min)。
(15) 用50μl含有去DNA酶的RNA酶A(20μg/ml)的TE重新溶解核酸,温和几秒钟振荡,贮存于-20℃。
2、质粒DNA的酶切、回收与连接
质粒DNA的酶切
(1) 提取的质粒DNA用相应的酶进行酶切。
酶切体系为:
单酶切双酶切
ddH2O 8μl ddH2O 8μl
质粒DNA 10μl 质粒DNA 10μl
10×buffer 2μl 10×buffer 2μl
限制性内切酶Ⅰ0.4μl 限制性内切酶Ⅱ0.4μl
限制性内切酶Ⅲ0.4μl 混匀,简单离心后,37℃水浴反应2-4h。
酶切产物进行1%琼脂糖凝胶电泳检测。
(2) 单酶切需要用小牛肠碱性磷酸酶(CIAP)去除质粒DNA两端的磷酸基,以防止自身环化。
①酶切产物纯化。
取酶切产物加1/10体积3mol/LNaAc和2倍体积预冷无水乙醇,冰上放置5min,12000rpm离心5min,回收沉淀,用预冷的70%酒精洗涤沉淀2次,倒置风干,用25μl TE Buffer溶解。
②CIAP的去磷酸反应。
DNA 25μl
ddH2O 19μl
10×AP buffer 5μl
CIAP 1μl
混匀,简单离心后,37℃反应1h。
③去磷酸产物纯化。
取去磷酸产物加1/10体积3mol/LNaAc和2倍体积预冷无水乙醇,冰上放置5min,12000rpm离心5min,弃上清,倒置风干,用20μl 以下的TE Buffer溶解。
质粒DNA的回收
采用TaKaRa Agarose Gel DNA Purification Kit Ver.2.0进行凝胶回收。
(1) 对酶切产物进行1%琼脂糖凝胶电泳。
在紫外灯下切出含有目的DNA 的琼脂糖凝胶,用纸巾吸尽凝胶表面的液体。
此时应注意尽量切除不含目的
DNA 部分的凝胶,尽量减小凝胶体积,提高DNA回收率。
(2) 切碎胶块。
称量胶块重量,以重量:体积1:3(1mg=3μl)向胶块中加入胶块融化液DR-I Buffer。
(3) 均匀混合后,75℃加热融化胶块,此时应间断振荡混合,使胶块充分融化(约6-10分钟)。
胶块一定要充分融化,否则严重影响DNA回收率。
(4) 向上述胶块融化液中加入DR-I Buffer量的1/2体积量的DR-II Buffer,均匀混合。
(5) 将试剂盒中的Spin Column安置于Collection Tube上。
(6) 将上述操作(4)的溶液转移至Spin Column中,12000 rpm离心1 min,将滤液再加入Spin Column中离心一次,可以提高DNA的回收率。
(7) 将500μl的Rinse A加入Spin Column中,12000 rpm离心30 s,弃滤液。
(8) 将700μl的Rinse B加入Spin Column中,12000 rpm离心30 s,弃滤液。
请确认Rinse B中已经加入了指定体积的100%乙醇。
(9) 重复操作步骤(8)。
(10) 将Spin Column安置于Collection Tube上,12000 rpm离心1 min。
(11) 将Spin Column安置于新的1.5 ml的离心管上,在Spin Column膜的中央处加入25μl的预热至60 ℃灭菌蒸馏水或Elution Buffer,室温静置1 min。
(12) 12000 rpm离心1 min洗脱DNA,4 ℃保存备用。
质粒DNA的连接
将酶切回收的目的片段DNA和载体DNA用T4 DNA ligase进行连接,连接产物可直接转化大肠杆菌。
连接体系:
PEG4000 1μl
Vector 0.5μl
DNA 5.5μl
10×ligase buffer 2μl
T4 DNA ligase 1μl
先将Vector和DNA混合好,置于45℃水浴5min,马上置于冰上冷却,再加入其他成分,混匀,简单离心后,置16℃过夜。
3、大肠杆菌感受态细胞的制备(CaC12法)
全过程都要在冰上进行,且动作尽量要轻
(1) 从37℃培养16-20h的平板中挑取单菌落,转到5ml LB液体培养基中,
37℃,250rpm,培养过夜(约16h)。
(2) 按1%接种量接种到含有50ml LB液体培养基的500ml三角瓶中,37℃,250rpm,培养2-2.5h(定时检测OD600值,使OD600≈0.4)。
(3) 将菌液转移到一个无菌的、一次性使用的、用冰预冷的50ml离心管中,冰上放置10min,使培养物冷却至0℃。
(4) 于4℃,4000rpm离心10min,以回收细胞。
(5) 倒出培养液,将管倒置1min以使最后的痕量培养液流尽。
(6) 每50ml初始培养液用30ml冰预冷的0.1mol/L CaC12溶液重悬细胞沉淀。
(7) 于4℃,4000rpm离心10min,以回收细胞。
(8) 倒出培养液,将管倒置1min以使最后的痕量培养液流尽。
(9) 每50ml初始培养液用2ml冰预冷的0.1mol/L CaC12溶液重悬细胞沉淀,置于冰上4-10min,分装成200μl /1.5mL离心管中,于-70℃保存备用。
4、大肠杆菌的转化
(1) 取DNA连接物10μl,冰浴下加入到200μl大肠杆菌感受态细胞中,混匀,冰上放置30min;其间可轻摇管2-3次,防止菌体沉于管底。
(2) 将管放入预加温至42℃的循环水浴中,热激90s,不要摇动管。
(3) 快速将管转移到冰浴中,室细胞冷却1-2min。
(4) 每管加入到800μlSOC液体培养基中,37℃,200rpm温育40min-1h,以使大肠杆菌复苏,并表达质粒编码的抗生素抗性标记基因。
(5) 将适当体积(每个90mm平板达200μl)已转化的感受态细胞转移到含相应抗生素的SOB平板上,室温放置直至液体被吸收。
(6) 倒置平皿,于37℃培养12-16h。
5、重组子的鉴定
(1) 重组子的PCR鉴定:
①从平板上挑取白色单菌落,于装有1ml LB(含有相应抗生素)的1.5ml离心管内37℃培养过夜;
②取1μl菌液作为模板,对单菌落进行PCR鉴定,初步筛选出含目的片段的阳性菌落。
(2) 重组质粒的酶切鉴定:
将待测阳性菌落进行扩培,用SDS碱裂解法进行质粒DNA的提取,通过酶切分析对阳性重组子进行最终确认。