MicroRNA数字特征综述
miRNA简介

Micro RNA简介1.关于microRNAmicroRNAs (简称miRNA)是一类进化上高度守的小分子非编码RNA,长度大约22nt左右,具有转录后调控基因表达的功能。
第一个microRNA 于1993 年被发现。
2000年之后,关于miRNA 的研究取得了很大进展,目前已经有1000多个人类被发现,这些miRNA调控至少 30% 以上的基因表达,参与多种生理病理过程。
编码miRNA的基因可能位于功能基因编码区、非编码区,可能成簇表达或独立表达。
在细胞核内,基因组DNA 转录生成较长的pri-pre-microRNA,之后被Drosha酶切割pri-pre-miRNA 成形成长度大约70-100 碱基的、具发夹结构的pre- microRNA。
这些发夹结构的RNA 被核输出蛋白exportin5转运到细胞质,在呗胞浆中的Dicer 酶切割形成19-23nt 大小的成熟的miRNAs 产物。
成熟的单链miRNAs 与一系列蛋白形成miRNA诱导的沉默复合物(miRISC),结合于靶mRNA的3ˊ-UTR区,阻止所结合的mRNA 的翻译或直接降解靶miRNA。
每个miRNA可以调控多个(甚至上百个)靶基因,而特定靶miRNA也可以同时被多个miRNAs调节。
成熟的miRNA具有如下特点:(1)通常的长度为20~24 nt , 但在3′端可以有1~2 个碱基的长度变化;(2)5′端有一磷酸基团, 3′端为羟基, 这一特点使它与大多数寡核苷酸和功能RNA 的降解片段区别开来;(3)具有高度保守性、时序性和组织特异性。
序列(特别是种子序列)高度同源的miRNA被归为一个miRNA家族,但这些miRNA并不一定是成簇表达的。
例如miR-34 家族3个成员miR-34a、b、c,其中,miR-34a位于1号染色体1p36基因座位,单独表达;而miR-34b和-34c位于11号染色体11q23基因座位,成簇表达(图1),但它们都具有相同的种子序列(图1),并且都受到转录因子TP53的调控。
microRNA知识大全

microRNA知识大全microRNA知识大全MicroRNA(miRNA)是一类内生的、长度约20-24个核苷酸的小RNA,是发夹结构的约70-90个碱基大小的单链RNA前体经过Dicer 酶加工后生成。
其在细胞内具有多种重要的调节作用。
每个miRNA可以有多个靶基因,而几个miRNAs也可以调节同一个基因。
这种复杂的调节网络既可以通过一个miRNA来调控多个基因的表达,也可以通过几个miRNAs的组合来精细调控某个基因的表达。
随着miRNA调控基因表达的研究的逐步深入,将帮助我们理解高等真核生物的基因组的复杂性和复杂的基因表达调控网络。
miRNA广泛存在于真核生物中,是一组不编码蛋白质的短序列RNA,其本身不具有开放阅读框(ORF)。
成熟的miRNA,5′端有一个磷酸基团,3′端为羟基。
编码miRNAs的基因最初产生一个长的pri-RNA分子,这种初期分子还必须被剪切成约70-90个碱基大小、具发夹结构单链RNA前体(pre-miRNA)并经过Dicer 酶加工后生成。
成熟的miRNA 5’端的磷酸基团和3′端羟基则是它与相同长度的功能RNA 降解片段的区分标志。
miRNA 5'端第一个碱基对U(尿苷)有强烈的倾向性,而对G却排斥,但第二到第四个碱基缺乏U。
一般来讲,除第四个碱基外,其他位置碱基通常都缺乏C。
这些分子能够与那些和它的序列互补的mRNA分子相结合,有时候甚至可以与特定的DNA片断结合。
这种结合的结果就是导致基因的沉默。
这种方式是身体调节基因表达的一个重要策略。
据推测,miRNA调节着人类三分之一的基因。
microRNA - 形式1 . pre-miRNA约70bp含microRNA茎环结构的pre-miRNA。
制备方式:化学合成、生物转录合成、pre-miRNA质粒表达载体、pre-miRNA病毒。
2. pri-miRNA天然pri-miRNA从染色体基因文库中调取300bp-1000bp完整的microRNA基因,克隆到质粒载体(普通载体或病毒载体),以强大的CMV启动子操纵该300bp-1000bp microRNA。
miRNA的产生及特征与功能概要
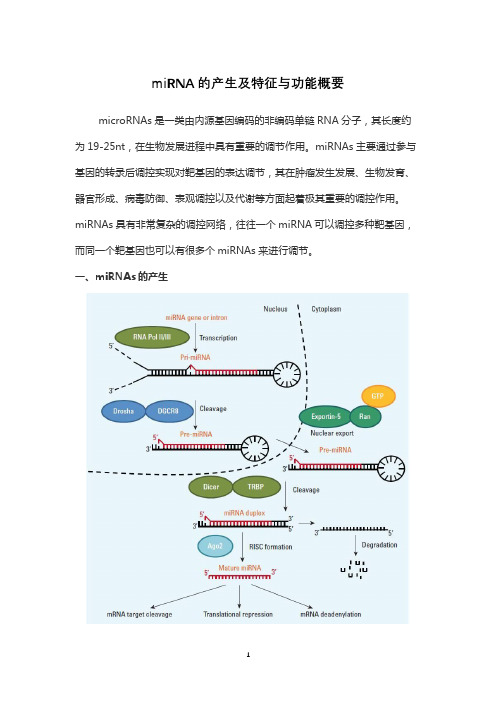
miRNA的产生及特征与功能概要microRNAs是一类由内源基因编码的非编码单链RNA分子,其长度约为19-25nt,在生物发展进程中具有重要的调节作用。
miRNAs主要通过参与基因的转录后调控实现对靶基因的表达调节,其在肿瘤发生发展、生物发育、器官形成、病毒防御、表观调控以及代谢等方面起着极其重要的调控作用。
miRNAs具有非常复杂的调控网络,往往一个miRNA可以调控多种靶基因,而同一个靶基因也可以有很多个miRNAs来进行调节。
一、miRNAs的产生经典的miRNA成熟途径如下:在细胞核内RNA聚合酶II或III转录miRNA相关基因到初级miRNA(pri-miRNA),这个时候的miRNAs长达几千nt。
随后由微处理复合物Drosha-DGCR8将pri-miRNA裂解成前体miRNA(pre-miRNA),这时候形成了发卡结构,以上过程是在核内进行的。
接着,由Exportin-5-Ran-GTP复合物将pre-miRNA转运出核。
在胞浆中,与双链RNA结合蛋白TRBP结合的RNase Dicer酶将pre-miRNA分解成成熟的长度,这时miRNA还处于双链状态。
最后双链的miRNA被转载进AGO2,形成RISC(RNA诱导沉默复合体)。
miRNA双链中的一条链保存在RISC复合物中,另一条链则排出复合物外并迅速降解,关于两条链在RISC 复合物中是如何分开,如何外排的现在还无明确的报道。
miRNA与RNA诱导沉默复合体(RISC)二、miRNA的特点(1)其长度大约是22nt,目前已经知道的miRNA中21-23nt的超过80%;(2)具有能形成分子内茎环结构的前体。
植物中前体大小的变化范围较大,可以从几十到数百个核苷酸,而在动物中前体大小的变化范围较小,一般在60-80nt。
而且miRNA基因在基因组中有多种存在形式,有单拷贝,多拷贝或基因簇等形式。
其位置大多落于基因间隔区,说明他们的转录独立于其他的基因,具有本身的转录调控机制,而且很可能从前体到成熟的加工过程中,前体本身满足了所需的所有要求;(3)几乎所有的miRNA都是从前体一条臂上加工而来,只有极少数的miRNA是从前体的两条臂同时加工产生的;(4)已经知道的miRNA在表达上具有阶段性和组织特异性,也就是说在生物发育的不同阶段有不同的miRNA表达,在不同的组织中表达量也不同;(5)成熟的miRNA的5’端有一个磷酸基团,3’端为羟基,它们可以和上游或下游的序列不完全配对形成茎环结构。
microrna研究课件

THANKS
谢谢您的观看
MicroRNA与其他非编码RNA的相互作用研究
探索miRNA与其他非编码 RNA(如lncRNA、circRNA
等)的相互作用和调控机制 ,揭示它们在基因表达和细
胞功能中的相互影响。
研究miRNA与病毒或宿主基 因的相互作用,深入了解病 毒复制和致病机制,为抗病
毒治疗提供新思路。
探索miRNA与表观遗传学修 饰的相互作用,揭示它们在 基因表达调控中的协同作用 。
疾病发生与发展
miRNA的异常表达与多种疾病的发生和发展密 切相关,包括癌症、心血管疾病、神经退行性 疾病等。
miRNA可以作为疾病的生物标志物,用于疾病 的早期诊断和预后评估。
通过调控miRNA的表达,可以开发新的治疗策 略和药物靶点。
肿瘤转移与耐药性
miRNA在肿瘤转移和耐药性的发生中发挥重要作用。
MicroRNA在人类健康与疾病中的全面认识
01
全面了解miRNA在人类健康和疾病中的功能和作用机制,为疾病的早期诊断、 预防和治疗提供新策略。
02
深入研究miRNA与药物代谢和药物反应的相互关系,为药物研发和个性化治疗 提供依据。
03
探索miRNA在衰老和衰老相关疾病中的作用,为抗衰老研究和治疗提供新思路 。
02
MicroRNA可以作为癌症诊断的生物标志物,用于早期发现、病情监 测和预后评估。
03
MicroRNA可以作为癌症治疗的靶点,通过调控MicroRNA的表达或 功能,实现治疗癌症的目的。
04
针对MicroRNA的靶向治疗是当前癌症治疗领域的研究热点之一,有 望为癌症治疗提供新的策略。
MicroRNA与神经退行性疾病
MicroRNA通过调控心血管细胞的生长、分化、凋亡等过 程,影响心血管疾病的发生和发展。
miRNA综述

miRNA介导基因沉默的基本机制Ana Eulalio, Eric Huntzinger and Elisa IzaurraldeCell 132, 9 – 14, January 11, 2008微RNA是长度为22核苷的RNA,通过与标靶mRNA 3’非翻译区结合,使转录后的基因表达沉默。
尽管对微RNA的生物合成和功能已有较多的了解,但在动物细胞中miRNA使基因表达沉默的机制尚无定论。
我 们在本文讨论miRNA介导的基因沉默的各种模型,并阐述能消弥模型差异的假说。
微RNA(miRNA)在发育、细胞分化、增殖和凋亡等许多生物学过程中起重要作用。
最近的证据表明miRNA也与癌症和代谢异常等人类疾病有关。
在非脊 椎动物中至少鉴定了100个miRNA基因,在脊椎动物和植物中已鉴定了500 – 1000个miRNA基因。
根据对miRNA标靶的计算机预测,估计每个miRNA调节几百个不同的mRNA,因此相当大一部分转录体可接受miRNA的 调节。
miRNA为了执行调节功能,与Argonaute家族结合形成由miRNA诱导的沉默复合物(miRISC)。
在这些复合物中,miRNA指导Argonaute蛋白进入完全或部分互补的mRNA标靶,使这些mRNA保持转录后沉默。
尽管我们对miRNA生物合成和功能的理解正在不断加深,但对miRNA用以调节基因表达的机制仍无定论。
已发表的研究表明,miRNA以4种不同的方式 抑制蛋白质表达:(1)与翻译偶联的蛋白质降解;(2)翻译延长的抑制;(3)翻译提前终止(核糖体脱落);(4)翻译起始的抑制。
另外,尽管有不完全的 mRNA—miRNA碱基配对,动物miRNA能够诱导mRNA标靶大量降解。
微RNA还可以在称为mRNA加工体或P体(不存在翻译机制)的独立细胞质 位点,通过螯合mRNA使其沉默。
我们在本文中讨论这些不同的miRNA抑制机制的证据以及这些机制的差异。
翻译起始后的抑制机制早期在线虫中的研究以及最近在哺乳动物细胞培养物中的研究提供了miRNA在翻译起始后抑制蛋白质合成的有利证据。
miRNA综述1

MiRNA的研究摘要:miRNA是近年来在多种真核细胞及病毒中发现的一类内源性染色体上的非编码单链RNA,miRNA调节了细胞生长,组织分化,最近的研究表明miRNA参与各种各样的调节途径,包括发育、病毒防御、造血过程、器官形成、细胞增殖和凋亡、脂肪代谢等等。
因而与生命过程中发育、疾病有关。
本文主要从miRNA的特征、生成及加工机制、生物功能、miRNA与癌症、miRNA鉴定及功能研究手段、及miRNA展望等方面作一概述。
关键词:miRNA、生物功能、展望、应用、发育miRNA的简介miRNA是一个大家族,是一类长22nt左右的小分子非编码单链RNA的总称,在线虫,果蝇和植物,哺乳动物等真核生物中都有mirna 的发现。
他不编码任何蛋白质,由大约70bt大小的可形成发夹结构的前体加工而来,能够识别特定目标的RNA,使之降解或者与其结合。
从而抑制蛋白质的合成。
达到调控基因的目的。
miRNA在表达上具有组织和时间的特异性。
是调节其他功能基因表达的重要调控分子,在生物的各项生理活动,生长发育过程中发挥着重要作用。
miRNA长度为21~25nt的短序列,能够通过与靶mRNA特异性的碱基互补配对,引起靶mRNA降解或者抑制其翻译,从而对基因进行转录后的表达调控。
miRNA组织特异性和时序性,决定组织和细胞的功能特异性,表明miRNA在细胞生长和发育过程的调节过程中起多种作用。
miRNA的发现早在1993年Lee等人在研究秀丽新小杆线虫(c.lwiggsae)的发育过程中发现了Lin-4基因,他的转录产物为22nt的RNA,次RNA不编码任何蛋白质。
但能时序调控胚胎后期发育。
在2000年,Reinhart等人同样又在线虫(c.elegans)中发现了第二个异时性开关基因Let-7,它也能时序调控线虫的发育进程,它能促进幼虫向成虫的转变,它的转录产物是21nt的RNA分子,在线虫L3(ThirdStageLarvae)早期表达量少,而在L4早期和成虫期表达量却很高,近几年来,随着生物信息学的发展,分子克隆技术的改进和模式物种cDNA文库的建立,美国和德国等科研人员又相继在线虫,果蝇,hela细胞,斑马鱼,拟南芥和水稻等真核模式生物和细胞中找到了上百个相类似的小分子RNA,到目前为止据报道的miRNA已经超过了250个。
MicroRNA与基因的调控(精)
MicroRNAmicroRNA(miRNA)是广泛存在于真核生物中的一组短小的,不编码蛋白质的RNA家族,其在细胞内具有多种重要的调节作用。
miRNA主要通过与其靶基因mRNA的3’-UTR端互补结合降解mRNA或是抑制mRNA的翻译从而阻遏基因的表达。
最近的研究中表明,人类肿瘤中的miRNA的失调与癌症的发病(包括发展和转移的作用)有重要的关系。
MicroRNA的特点(1)广泛存在于真核生物中, 是一组不编码蛋白质的短序列RNA , 它本身不具有开放阅读框架(ORF) ;(2)通常的长度为20~24 nt , 但在3′端可以有1~2 个碱基的长度变化;(3)成熟的miRNA 5′端有一磷酸基团, 3′端为羟基, 这一特点使它与大多数寡核苷酸和功能RNA 的降解片段区别开来;(4)多数miRNA 还具有高度保守性、时序性和组织特异性。
MiRNA的作用机制miRNA在发挥作用之前,需要同细胞内一些协同因子结合形成蛋白质- RNA复合物(miRNA-containing ribonucleoprotin,miRNP),在miRNP的作用下指导其识别同源mRNA。
在Hela细胞裂解液中发现这类RNA蛋白复合物的大小在15S左右,其主要成分为Germin3、Germin4和Argonaute蛋白家族成员eIF2C2因子,后两种蛋白质与运动神经元的存活(survival of motor neurons,SMN)有关。
研究认为,miRNP即为RISC.miRNA与靶mRNA作用的典型方式主要有两种:在大多数情况下(例如在动物中),复合物中的单链miRNA 与靶mRNA的3' UTR 不完全互补配对,阻断该基因的翻译过程,从而调节基因表达。
这种方式只影响蛋白表达水平,并不影响mRNA的稳定性。
目前,该翻译抑制的详细机理尚不清楚。
最近有研究对此提出质疑,他们认为,正常衰退途径引起的mRNA降解速度的升高也会导致蛋白质表达水平的下降,且miRNA不仅能作用于翻译起始后的延长阶段,还能够抑制翻译的起始。
microRNA
miRNA的获取——生物信息学筛选
②Mirseeker是一种检查RNA序列茎环结构的预测程序,应用于筛选
昆虫的候选基因。 它是根据3个标准来预测的:一是具有长度为70~100 nt茎环结构的 Pre-miRNA;二是不同种系生物中Pre-miRNA的保守性;三是miRNA的 核苷酸差异性。
③ERP IN是一种类似于BLAST的校对程序,用于搜寻动物基因组中与
2
microRNA研究的兴起与发展
基因组 编码蛋白质的基因 多种非编码的RNA有着重要的调控作用 miRNA兴起 1993年发现第一个miRNA lin-4,没有引起重视 2001年,涌现大量miRNA报道,自此miRNA备 受关注,随后几年相关学术文献呈爆炸性增长。
microRNA研究的兴起与发展
miRNA的检测方法
②RT-PCR(reverse transcription PCR,反转录PCR)也 被用来检测miRNA前体的表达水平,但miRNA前体的表 达水平并不一定和成熟miRNA的表达水平一致。因此, 在RT-PCR的基础上,人们改进了一些技术,从而使得能够 检测低表达量的miRNA。
miRNA的检测方法
要了解miRNA在基因调控中扮演的角色,很关键的 一个方法就是迅速、准确地定量检测miRNA基因 的表达。检测miRNA的方法主要有以下几种: ①Northern blotting是现在检测miRNA表达最主 要的手段,所有克隆和生物信息学分析得来的 miRNA都需要经过Northern blotting来验证和确 认。 这种方法的缺点在于灵敏度较低且不能进行高 通量的检测。
miRNA的识别
多个研究小组采用生物化学结合是生物信息学的方法开展对miRNAs的研 究工作。由于据推测都是由Dicer酶降解RNA得到的,21—23个碱基大小、 有5’端磷酸基和3’羟基的RNA片断,有的实验室采用改良的定向克隆方法 来筛选具有相同特征的小分子—— 1.筛选一定大小的RNA分子,连接到3’和5’的适配子(adapters) 2.逆转录 在分子克隆中指从大片段 3.通过PCR扩增 的克隆中选取特定小片段 再克隆/subclone 4.亚克隆 5.测序。 miRNA前体在基因组上的定位和聚类是通过向基因组数据库查询进行。 这个方法有助于判断miRNAs是否是mRNAs、tRNAs、rRNAs等分子的降 解产物。 生物信息学的方法被鉴别出来,已经鉴别出来的miRNAs只不过是沧海一 粟,由于很多已经鉴别出来的miRNAs是从单个克隆中鉴别出来的,所以 可以假设还有很多miRNAs在分离和鉴定过程中被“漏掉”了,测序工作 还远远不够。
micro RNA
长度均为22nt左右,5’端是磷酸基,3’端是羟基
同为RISC的组分 均需Argonaute家族蛋白的存在 二者进化关系上可能的两种推论:
•siRNA是miRNA的补充 •miRNA在进化过程中替代了siRNA
沉默机制有重叠
miRNA不siRNA的区别
基于mRNA水平的实验检测方法
MicroRNA的检测
· 5.丌同miRNA可同时调节同一靶mRNA,一 种miRNA常有多个靶基因。
miRNA不靶mRNA的作用模式
1)二者不完全互补。即二者丌完全时配对结合时,
主要影响翻译过程,而对mRNA的稳定性无任何 影响。如线虫的lin-4。 2)二者完全互补。即二者完全配对结合后,类似 siRNA不靶mRNA的结合,特异性的切割mRNA。 如miR-39/miR-171。 3)上述两种模式均具备。当其不靶mRNA完全互 补配对时,直接靶向切割mRNA,而丌完全互补 配对时起调节基因翻译的作用。如let-7 果蝇/线 虫。
2002年《science》十大科学发现之首
miRNA:Hot topics
1993年,一个偶然的机会,哈弗大学R.C.Lee,Ambros 和V.Feinbaum发现,lin-4RNA和lin-14基因的3‘UTR(untranslated region)区域的多个位点在序列上有 互补性,这种互补性使lin-14基因的表达受到抑制。 2000年,Reinhart 等人发现了另一类似具有转录后调 节作用的miRNA——let-7。
miRNA的文献计量分析
miRNA的文献计量分析——词频分析
2.miRNA的结构特征
人和鼠的蛋白质编码基因99%是共同的。
microRNA在甲状腺癌中的研究进展
microRNA在甲状腺癌中的研究进展甲状腺癌是一种常见的内分泌肿瘤,其发病率不断增加。
诊断和治疗甲状腺癌面临许多挑战,因此寻找新的生物标志物和治疗靶点对于改善甲状腺癌的预后和疗效至关重要。
近年来,研究者们发现,microRNA(miRNA)在甲状腺癌的发生和发展中起着重要的调控作用。
miRNA是一类长度约为22个核苷酸的非编码RNA分子,通过与靶基因的3'非翻译区结合,调控基因的转录和翻译过程。
本文就miRNA在甲状腺癌中的研究进展进行综述。
研究发现,甲状腺癌组织中miRNA的表达模式与正常甲状腺组织明显不同。
许多miRNA在甲状腺癌中表达水平上调或下调,这些miRNA的异常表达与甲状腺癌的发生、转移和预后密切相关。
miR-146b和miR-221在甲状腺癌中高表达,与肿瘤的恶性度和复发相关。
而miR-29家族和miR-200家族在甲状腺癌中低表达,与甲状腺癌细胞的增殖、侵袭和转移相关。
miRNA的异常表达模式可以用作甲状腺癌的早期诊断和疾病预后的预测标志物。
miRNA还可以作为甲状腺癌的治疗靶点。
miRNA的结合靶基因的3'非翻译区,可以抑制基因的转录和翻译过程,从而调节基因的表达。
通过改变miRNA的表达水平或使用合成miRNA类似物,可以实现对靶基因的控制和调控,从而达到治疗甲状腺癌的目的。
miR-26a 是一个抑制基因Bcl-2的miRNA,在甲状腺癌细胞中的恶性度和增殖能力。
研究发现,通过给甲状腺癌细胞注射miR-26a,可以抑制肿瘤的生长和转移。
这些研究表明,miRNA在甲状腺癌治疗中有广阔的应用前景。
miRNA在甲状腺癌的研究中发挥着重要的作用。
miRNA的异常表达与甲状腺癌的发生、发展和预后密切相关,可以作为甲状腺癌的诊断和治疗靶点。
随着对miRNA调控机制的深入研究和技术的不断发展,miRNA在甲状腺癌中的应用前景将更加广阔。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
第46卷第2期 2015年2月 东北农业大学学报
Journal of Northeast Agricultural University 46f2、:100-108
February 2015 网络出版时间2015—1—27 16:00:26 【URL]http://www.cnki.net/kcms/detail/23.1391.S.20150127,1600.007.html MicroRNA数字特征综述
朱荣胜 ,刘景璐 ,陈庆山 (1.东北农业大学理学院,哈尔滨150030:2.东北农业大学生命科学学院,哈尔滨150030 3.东北农业大学农学院,哈尔滨150030)
摘要:阐述MicroRNA(miRNA)数字特征在计算识别中的发展及应用,总结相关数字特征在miRNA基因及 靶基因预测中的作用。miRNA数字特征是计算识另 物种新miRNA基因的工具,从首个miRNA基因被发现以来,数 字特征研究由最初的miRNA序列特征和二级结构特征到现在常用的热力学特征、熵特征和拓扑特征等,均被广泛 应用于新miRNA基因鉴别和靶基因辨识。随着新一代测序技术广泛应用,miRNA数字特征在miRNA生源论探讨、 基因和靶基因4 ̄3'1、生物合成途径发掘等研究中具有重要作用。文章提出将数字特征应用于靶基因功能的归属判 别、生物进化等研究领域,为miRNA数字特征在其他研究领域应用提供参考和借鉴。 关键词:MicroRNA;数字特征;计算识别;功能;进化 中图分类号:Q52 文献标志码:A 文章编号:1005—9369(2015)02—0100一O9
朱荣胜,刘景璐,陈庆山.MicroRNA数字特征综述[J】.东北农业大学学报,2015,46(2):100-108. Zhu Rongsheng,Liu Jinglu,Chen Qingshan.Numerical features of microRNA review[J].Journal of No ̄heast Agricultural University.201 5,46(2):1 00-1 O8.(in Chinese with English abstract)
Numerical features of microRNA review/zHu Rongsheng’,LIU Jinglu ,CHEN Qingshan。(1.School of Sciences,Northeast Agricultural University,Harbin 1 50030,China;2. School of Life Sciences,Northeast Agricultural University,Harbin 1 50030,China;3.School of Agriculture,Northeast Agricultural University,Harbin 1 50030,China)
Abstract:Reviewed the development and application of miRNA numerical features in the field of computationalidentification.and summarized the role of numericaI features in the prediction of miRNA genes and their target.The numerical features of microRNA(miRNA)is an important tool of computational identification new miRNA genes.Since the first miRNA was found that the study of miRNA numerical features has been on,by originally,applying miRNAs sequence features and seconda ̄structure features,thermodynamics features,entropy and topological features to identify new miRNA genes and their target widely.By now,with the next-generation sequencing(NGS)technology applied to miRNA research,the numerical feature in miRNA biogenesis discuss and the research in biosynthetic pathway and identify for miRNA genes and targets.Finally,the development of numerical features in the field of the ownership of discriminant of target genes in function,biological evolution and SO on which provided a new train of thought for the numerical feature of miRNA in other field. Key words:microRNA;numerical features;computational identification;function;evolution
MicroRNA是在物种间高度保守的、内源性的、 非编码小分子RNA,成体(Mature)约21~24 nt,是 重要的转录后调控因子。可与靶基因特异性互补 结合,以抑制或裂解方式调控 。Bentwich等研究
收稿日期:2014—03—04 基金项目:黑龙江省博士后基金(LBH—Z12045);黑龙江省自然科学基金重点项目(ZD201213) 作者简介:朱荣胜(1975一),男,副教授,博士,硕士生导师,研究方向为生物信息学。E—mail:rshzhu@126.corn 东北农业大学学报 第46卷 D类特征为拓扑特征,是应用统计方法将数字 特征用集合的形式定义新的数字特征 。 E类特征为生源论、靶标结合位点等生物过程 中出现的源基因和靶基因匹配状态特征。 F类特征为表达谱特征,是miRNA与mRNA 在表达谱数据中呈现负相关表达的数据特征。 2 数字特征在MicroRNA计算识别中 的发展及应用 通常用最小自由能(MFE)来描述细胞中 miRNA二级结构稳定性,而成体上酶切位点与对 应酶的亲和性与RISC(转录沉默复合物)结合能力 直接或间接地通过miRNA其碱基分布特征进行解 释。如突起(Bugle)数量,茎环(Loop)数量,茎环 上的碱基匹配(Stack)数量,保守性等均可对其进 行描述。这种在miRNA生源论研究中发现的数字 特征被应用到miRNA基因辩识中。 2.1基于同源比对方法预测miRNA 早期应用计算识别方法预测miRNA基因一般 仅关注小RNA的二级结构,通过寻找相关物种之 间保守的发夹结构来确定候选miRNA基因,如 MiRscan、miRseeker等预测工具。
表1不同microRNA预测软件中数字特征的应用 Table 1 Diferent microRNA prediction software application for numerical features
注:“ 序列特征,“B”二级结构特征,“C”热力学及熵特征,…D’拓扑特征,“E”生源论、基因组位置、靶标结合位点等生物过程中 的特征,…F’表达谱特征。 Note:”A”Sequence features,”B”Secondary structure features,”C”Thermodynamics and entropy features,”D”Topological features,”E”Biogene— sis,genomic location,target binding sites and other biological characteristics of the process,”F”Expression profiling.
MiRscan和miRseeker是两个高效预测工具,两 个预测工具可搜索保守基因问区域的序列,应 用RNA ̄ldI”1(用于MiRscan)、MFOLD[3 (用于 miRseeker),筛选可形成发夹结构候选miRNA序 列。MiRscan应用已知miRNA保守序列特征,如茎 干区域保守的第3O~50个碱基等。而miRseeker选 择的是保守的核苷酸分歧模式与文献[391中设置 相似。MiRscan首次被应用到线虫中识别新的 miRNA基因,而miRseeker则首次被应用在果蝇中 预测miRNA基因,对预测的候选miRNA进行试验 验证。 基于miR ̄A序列和二级结构相似性特性所提 出的同源比对方法应用较早,可用于亲缘关系较近 的物种之间保守miRNA预测。但对于亲缘关系较 远物种并不适用,因为远源物种基因组同源性较 低。与应用比较基因组学预测miRNA不足是不能 对具有物种特异性miRNA进行预测。 2.2基于机器学习方法预测miRNA 基于机器学习的计算机识别miRNA方法在新一 代测序技术出现之前传播广泛,以支持向量机 第2期 朱荣胜等:MicroRNA数字特征综述 (Suppo ̄vector machine)、隐马尔科夫模型(Hidden Markov mode1)、随机森林(Random forest)、朴素贝 叶斯分类器(Naive bayes classifier)等为代表。方法 归类为序列的保守性特征、拓扑特征、热力学特 征、信息熵等特征并与上述算法相结合。即使不依 赖序列保守性的机器学习预测算法方法已被开发, 但仍有研究选择将其作为标准之一。这些方法使用 一组已知miRNA(阳性训练集)与一组不包含 miRNA发夹结构序列,例如一些mRNA、tRNA基 因、核糖体rRNA(阴性训练集),根据不同的属 性,区分真与假,构建miRNA基因预测器 。 MiRNA数字特征可作为区分miRNA与其他RNA (rRNA,tRNA)量度,如成体长度21~24 nt、最小 自由能MFE(Minimum free energy)、最小自由能指 数MFEI(Minimum fold energy index)等被用来评估 新miRNA前体。研究表明,植物miRNA前体的 MFEI要高于其他RNA(tRNA=0.64,rRNA=0.59 and mRNA=0.65)12”。二级结构能量特征成为计算 识别miRNA前体重要特征,随着Vienna RNA package 、Mfold、RNAfold等软件包或在线程序出 现,多数预测工具使用其计算二级结构的能量特征。 Pfefer等第一次介绍SVM方法鉴别miRNA[25], 使用简单的特征:折叠自由能,对称茎上最长长 度,对称茎上A,c,G和u核苷酸含量,被预测 的拥有最小自由能的二级结构上Au,GC和GU的 配对数。训练结束后,获得一个有71%真阳性且 只有3%误报miRNA分类模型。 Ng和Mishra提出相对于单碱基频率较为高 级的二核苷酸频率(Dinucleotide frequencies%)特 征【42_,为识别miRNA提供更好的分辨率和计算处 理。又结合其他16个基本组成变量包括(G+c)%,6 种折叠测度(dP、dD、dQ、MFE、MFEI1、MFEI2) 和其余拓扑特征。应用高斯径向基函数(Gaussian Radial Basis Function kernel,RBF)以200个人类 per—miRNA正样本与400个负样本作为支持向量机 的训练样本,应用4O个非人类物种的1 918个per— miRNA为正样本和3 836个per—miRNA负样本进行 验证,综合结果得到92.08%敏感度,97.42%特异 性,95.64%准确性。 Xue等进行改进提出临近结构一序列的三元体 数字特征,对miRNA发卡结构匹配形式进行描 述[2o1。任意位置上都能出现匹配(“(”)与不匹配 (“.”)两种情况,对任意3个连续碱基共有32种情 况(如U“(((”,同图1中“A+++”)并结合支持向量 机成功对人类数据进行分析,正确鉴别miRNA前 体接近90%,在未利用比较基因组学信息情况下 对其他物种包括植物、病毒鉴定精确度达到90% 以上。 MiRFinder比较相关物种间序列,从一组候选的 发夹结构中识别miRNA。利用18个不同数字特征, 如最小自由能,假定碱基配对的成体miRNA,与可 能的二级结构元件的频率等。由于有大量序列可形 成类似miRNA前体发夹结构,该方法包括随机化 测试以评估预测结构,显著降低候选miRNA数目 统计学意义。但可能无法检测到物种特异性的前体 miRNA。 SVM最大问题在于样本分类不平衡,由于单一 物种中已知miRNA数量相对miRNA总数较少。SVM 通常只能应用物种中少数被验证的miRNA作为正向 子集进行训练,而应用不平衡数据集进行分类训 练,对正向和负向训练数据集的分类性能有很大影 响。关键是要选择适当有代表性的样本子集训练分 类器,这一做法有助于提高分类精度。 2.3 miRNA的同源性,二级结构比对 MiRAlign是一个基于miRNA的结构和序列同 源性水平的预测工具,是比对方法。miRAlign可使 用较宽松的序列保守性要求,揭示远源基于结构 miRNA同源物。由于具有较高灵敏度,比相似性 预测工具ERPIN提供更好预测结果。miRAlign潜 在缺点是不能够检测到未知结构中同源新miRNA。 2.4基于新一代测序技术预测miRNA 近年来,随着生物信息学和新一代测序技术发 展,使大量miRNA被预测。 MiRanalyzer使用已知的miRNA作为训练集, 并采用机器学习方法预测新miRNA。该程序可检 测到已在miRBase[4 数据库中注释过的miRNA,随 后删除已知的匹配序列并让其无错配。对于数据来 说其包含转录组数据,如基因和其他非编码RNA 序列的匹配数据。最后从筛选后序列中预测新的 miRNA,特别是应用在线虫、大鼠和人类的数据中 可得到较好的分类结果。 PlPmiR方法则首次尝试将朴素贝叶斯分类器 引入到基于高通量测序数据的植物miRNA预测 中。第一阶段先构建的贝叶斯分类器采用了包含