L_苏氨酸发酵过程中乙酸的控制
发酵过程中pH值调控方法浅析

发酵过程中pH值调控方法浅析微生物发酵是一个十分复杂的生物化学反应过程,由于生物体的生长、繁殖、代谢有许多不确定性,并受环境影响比较大,而微生物细胞内同时进行着成千上万种不同的生化反应,各有各的调控机制,相互促进相互制约,因此,其控制过程也比较难,只能依据宏观现象或借助一些检测手段来推测微观变化,而后施行进行人为干预,进而引导其朝着我们设定的方向发展。
通过取样分析获得与微生物发酵相关的参数包括:物理、化学、生物三类。
(宏观现象不在本文赘述)。
1、物理参数包括:温度、罐压、搅拌转速、搅拌功率、空气流量、发酵液粘度;2、化学参数包括:酸碱度、基质浓度、溶解氧浓度、氧化还原电位、中间体物质浓度、产物浓度、比生产速率、尾气氧含量、尾气二氧化碳含量;3、生物参数包括:菌体浓度、比生长速率、菌体形态;各参数调控方式以及各参数之间错综复杂相互关系,今天不做展开,仅对pH值的进行详细分解。
第一部分现行的pH控制理论第一节为什么要控制pH值?1. 影响菌体生长:每种微生物都有其适合生长的pH值范围,原因有二:(1)对营养成分的影响:不同的pH值条件下,培养基中各营养物质的稳定性不同;各营养物质之间发生化学反应不同;有机碳、氮源的水解或变性情况不同。
进而影响菌体对营养物质的利用。
(2)对菌体生物酶活性影响:培养基中的H+或OH-直接影响胞外生物酶的活性,进而影响菌体生长。
(3)对菌体细胞结构的影响。
(4)对菌体细胞膜荷电影响:引起细胞膜的通透性发生改变,因而影响菌体对营养物质的吸收和代谢物排泄。
2. 影响菌体代谢:培养基中的H+、OH-首先作用在胞外的弱酸(或弱碱)上,使之成为易于透过细胞膜的分子状态的弱酸(弱碱),它们进入细胞后,再进行解离,产生H+或OH-离子,改变胞内原先的中性状态,影响酶的结构和活性。
进而影响菌体代谢。
3. 影响产物稳定性:不同的物质有不同的稳定pH值范围。
特别是胞外表达的产物,直接裸露于培养液中,给予合适的pH条件至关重要。
L_苏氨酸的生产方法及研究进展

第28卷第5期河南工业大学学报(自然科学版)Vo.l 28,N o .52007年10月Journa l o fH enan Un iversity of Techno l o gy(N atural Sc i e nce Edition)O c.t 2007收稿日期:2007 06 05基金项目:国家863项目(2006AA02Z216)作者简介:黄金(1979 ),男,安徽利辛县人,博士研究生,主要从事代谢调控方面的研究.*通讯作者文章编号:1673 2383(2007)05 0088 05L-苏氨酸的生产方法及研究进展黄 金,徐庆阳,陈 宁*(天津科技大学天津市工业微生物重点实验室,天津300222)摘要:介绍了L-苏氨酸的理化性质、用途、生产方法,重点介绍了发酵法生产L-苏氨酸的生产现状.依据代谢调控理论综述了L-苏氨酸生物合成调控机制及研究实例,并对L -苏氨酸市场前景进行了展望.关键词:L-苏氨酸;生产方法;研究进展中图分类号:TS201 2 文献标识码:A0 L-苏氨酸的理化性质苏氨酸(Threon i n e)化学名称为 -氨基- -羟基丁酸,1935年由W.C .Rose 在纤维蛋白水解物中分离和鉴定出来.1936年,M eger 对它的空间结构进行了研究,因结构与苏糖相似,故将其命名为苏氨酸[1].苏氨酸有4种异构体,天然存在并对机体有生理作用的是L-苏氨酸[2].其化学结构式为:图1 L-苏氨酸结构式分子式为C 4H 9NO 3,相对分子质量为119.12,熔点为255~256 ,比旋光度[ ]20D=-25.0~-29.0 (5m ol/L H C l),白色晶体或结晶性粉末微甜;解离常数p K COO H =2.15,p K N H2=9.12,等电点p I(25 )=5.64,溶解度为20g /100m L 水(25 ),不溶于乙醇、乙醚、氯仿等有机溶剂.1 L-苏氨酸的应用在人体和动物所需的8种必需氨基酸中,苏氨酸是仅次于蛋氨酸、赖氨酸、色氨酸的第4种氨基酸[3-6].苏氨酸在人的生长发育中发挥越来越重要的作用,被广泛应用于食品工业、饲料工业、以及医疗等方面.在食品方面,由于当前食品加工度高、生产量大、保存期长、流通领域广,营养素容易受到破坏和损失,需要加以补充.因此,食品强化剂在食品工业中的重要性日益突出,氨基酸类强化剂在食品强化剂生产中占有重要的地位.苏氨酸与葡萄糖共热可产生焦香味,有增香作用.一般与其他氨基酸合并用于强化谷物,有时也用于糕点、婴儿食品、牛奶中,起抗氧化作用[7].在饲料添加剂方面,苏氨酸和赖氨酸、蛋氨酸、色氨酸一起列为4大饲料添加剂,主要用于未成年仔猪和家禽等.苏氨酸是畜禽饲料中的重要元素,在畜禽饲料中添加苏氨酸,能提高饲料蛋白质的生物学价值,平衡各种氨基酸,促进蛋白质沉积,降低动物氨的排泄,减轻环境污染,改善饲养条件.在医药工业方面苏氨酸应用也十分广泛,其铁盐可作抗贫血药,可用于配置氨基酸输液和综合氨基酸制剂,同时其自身还具有促进人体发育和抗脂肪肝的药用疗效,也是制造高效抗生素单酰胺菌素的中间体,以及用于生化研究.在临床上苏氨酸还具有提高免疫的功能:苏氨酸有促进骨髓T 淋巴细胞前体分化发育成为成熟T 淋巴细第5期黄 金等:L-苏氨酸的生产方法及研究进展89胞的作用.苏氨酸是谷蛋白中第二限制性氨基酸,它的分子结构中含有羟基,对人体皮肤有保湿作用.在膜蛋白中苏氨酸的羟基能与寡糖链结合,对保护细胞膜起重要作用.此外,苏氨酸代谢生成甘氨酸,提供一碳单位.另外,苏氨酸是11种、14种、17种、18种、20种氨基酸大输液的主要成分之一.氨基酸大输液常用于手术前后、创伤、烧伤、骨折、营养不良、慢性消耗性疾病等的辅助治疗,在临床方面有着广泛的应用.2 L-苏氨酸生产方法研究进展L-苏氨酸的生产方法有蛋白质水解法[8]、化学合成法[9]和直接发酵法[10].蛋白质水解法和化学合成法因其存在种种弊端,工业化生产已经基本不再使用.直接发酵法以其生产成本低、资源节约、环境污染小等优点逐渐成为工业化生产L -苏氨酸的主要方式.2.1 蛋白质水解提取法苏氨酸主要以L 型存在于血粉、角蹄、玉米麸质粉、棉籽饼、丝胶等天然蛋白质中,可将上述蛋白质经水洗、搅碎、干燥、酸解得水解液经浓缩得浓缩液,用活性炭脱色、上柱,接收液作纸层析,收集苏氨酸部分.但该方法存在不少问题,操作复杂,需处理大量洗脱液,最主要的是苏氨酸在这些蛋白质中含量较低,尤其是提取目标氨基酸时,其他氨基酸作为废料处理掉,造成资源的浪费.2.2 化学合成法苏氨酸的化学合成路线,主要有巴豆酸法、乙酰乙酸乙酯法、甘氨酸铜法及不对称环氧化法等,其中路线较短、收率较高、具有较高工业价值的方法是甘氨酸铜法.2.2.1 巴豆酸合成法巴豆酸在乙醇中进行汞化反应,再经溴化、脱汞、氨化得DL-苏氨酸.该法反应条件缓和、产率也不低,但步骤较繁、溶剂成本高,加之含汞废液处理困难等,使其利用价值不大.2.2.2 乙酰乙酸乙酯合成法将乙酰乙酸乙酯氨化、还原,即可得DL-苏氨酸,该路线反应条件要求高,且产率低.2.2.3 甘氨酸铜合成法甘氨酸由于与Cu 2+络合,生成螯合物,从而增强了 氢的酸性,更利于其与乙醛缩合,最终脱去铜,生成DL-苏氨酸.Cu 2+起到了催化剂的作用.也有用五水硫酸铜或氯化铜与甘氨酸和乙醛来制备的.Aune 则采用中间体双(2,5-二甲基-4-恶唑烷羧酸酯)二水合铜,与甘氨酸反应合成DL-苏氨酸.这样,可以较大地提高产率.但成本也大大增加了.通常,采用廉价的Cu(OH )2CO 3和CuC l 2为催化剂.合成路线如图2.图2 甘氨酸铜合成路线2.2.4 不对称环氧化法不对称环氧化法,具有选择性高的优点,但由于其反应过程复杂,原料成本太高,难以实现工业化生产.该法以C H 2C HC H (OH )C H 3为原料,双键经环氧化后,与phCONCO 反应,生成五元杂环化合物,再酸化开环,生成DL-苏氨酸.2.2.5 非水相反应合成以苯为溶剂,将氰基乙酸丁酯与乙醛,在30~35 搅拌反应6h ,经过滤,酸化滤液,可得88%的苏氨酸.上述几种方法,各有优劣,其中甘氨酸铜合成法得到普遍使用.该法成本低,合成路线短,产率高(以甘氨酸计,可达80%以上).但铜盐的回收再利用,离子交换脱铜过程繁杂、且洗脱液不易浓缩.2.3 微生物发酵法直接发酵法是借助于微生物具有合成自身所需要的氨基酸的能力,通过对特定微生物的诱变处理,选育出营养缺陷型或结构类似物抗性菌株,以解除代谢调控中的反馈抑制和反馈阻遏,从而达到过量积累某种氨基酸的目的[11].随着基因工程技术的发展、工业微生物生物化学信息的增多、特别是工业生物载体系统的成功构建,从上世纪70年代末起前苏联研究者开始用基因工程技术构建苏氨酸菌种[6],为优良的L -苏氨酸生产菌株的筛选和产酸水平的提高提供了可靠的技术保障,使微生物直接发酵法生产L -苏氨酸成为一种廉价的工业化生产方法.90 河南工业大学学报(自然科学版)第28卷2.4 L -苏氨酸代谢调控育种研究2.4.1 L-苏氨酸合成及调控机制由L-天冬氨酸为起点产生L-苏氨酸和L -异亮氨酸的生物合成途径如图3所示.图3 L-苏氨酸生物合成途径及调控机制 整个合成途径中含有两大节点,即由L-丁氨醛酸合成L-赖氨酸或高丝氨酸以及由高丝氨酸合成L-甲硫氨酸或L-苏氨酸直至L-异亮氨酸.前一节点两条途径的代谢流的流向由高丝氨酸脱氢酶(H D)和二氢吡啶二羧酸合成酶对其共同底物丁醛氨酸的相对亲和性控制.这两个酶的编码基因分别为ho m 和dapA.在正常情况下,HD 的第五亲和性以及转化率是其竞争对手二氢吡啶二羧酸合成酶的25倍,因此碳源主要流向苏氨酸-甲硫氨酸生物合成途径.但当生产菌细胞内L-苏氨酸大量积累时,由于HD 对L-苏氨酸的变构抑制作用极为敏感,从而导致丁氨醛酸优先合成赖氨酸[12-15].L-苏氨酸的代谢调控机制中二氨基庚二酸、赖氨酸、甲硫氨酸、苏氨酸及异亮氨酸中的碳原子都来源于天冬氨酸.其共同途径的第一步是由3种不同的门冬氨酸激酶所催化的.天冬氨酸激酶I 能被苏氨酸抑制,它的合成被苏氨酸及异亮氨酸所阻遏.天冬氨酸激酶II 的合成则被甲硫氨酸所阻遏.而赖氨酸则能抑制天冬氨酸激酶!的活性,并且阻遏其合成.从丁氨醛酸到高丝氨酸的则由两种不同的高丝氨酸脱氢酶所催化的,高丝氨酸脱氢酶I 的合成可被苏氨酸及异亮氨酸阻遏,其中苏氨酸又能抑制其活性;而高丝氨酸脱氢酶I 的合成由甲硫氨酸所阻遏[16-17].2.4.2 育种实例20世纪50年代日本的志村、植村两位教授采用添加前体物的方法发酵生产苏氨酸.国外从60年代就开始有直接发酵法生产L-苏氨酸的报道.70年代末苏联的研究者们用基因工程菌规模生产L-苏氨酸.国内从1982年才开始有L -苏氨酸产生菌选育的报道,黄和容等报道了以钝齿棒杆菌(C orynebacteri u m crenatum )为出发菌株选育苏氨酸产生菌,产量为13g /L ;1986年檀耀辉等报道以乳糖发酵短杆菌(B revibacterium lacto fer m entum )出发选育出一株L-苏氨酸产生菌,在适宜条件下发酵72h 可产酸16g /L ;1990年沈阳药学院的常尊学等用亚硝基胍(NTG )和紫外线(UV )诱变处理黄色短杆菌获得一株苏氨酸高产菌株ME7(AHV rAEC rM et E th s),在含有葡萄糖10%的培养基中发酵48h 可积累L -苏氨酸17.9g /L.2000年韩国K.-H.Song H.-H.Lee 等构建一耐高盐浓度菌株H S528在5L 罐发酵产酸可达74g /L .2002年华东理工大学沈琼构建苏氨酸基因工程菌E.coli V NBk B -3507发酵48h 产酸52.7g /L .2005年广东肇庆星湖生物科技公司的王焕章、吴新等人利用大肠杆菌K12通过同源重组缺失苏氨酸脱氨酶基因,筛选L-Ile 营养缺失菌株,导入苏氨酸操纵子质粒,获得基因工程菌E.coliTHR 6,发酵32h 产酸可达75g /L .2006年韩国的M an -H yo ,H ong -W eon Lee 等利用蛋氨酸缺陷菌大肠杆菌MT201,添加生物素和蛋氨酸发酵33h 产酸可达80.2g /L .3 L-苏氨酸市场概况及展望苏氨酸为一种重要的氨基酸,主要用于医药、食品强化剂、饲料添加剂等方面.近年来,国内外市场对苏氨酸的需求逐年强劲增长,国际市场年增长率达到20%多.我国市场多年来对苏氨酸的需求平衡,近几年市场开始升温.国内生产不能满足市场需求,每年都需大批进口.预计今后国内外市场对苏氨酸的需求量还将以较大比率上升,短期内国内市场供需矛盾无法根本解决,仍需进口.因此,开发苏氨酸市场前景看好.最近几年,全球苏氨酸市场以每年20%多的增长率高速增长,亚洲、北美等地区的需求快速增长.需求的旺盛拉动了生产,1993年,全世界苏氨产量为4000,t 1996年为5000,t 1999年猛增至2.5万,t 2001年达3万,t 2002年达4万,t 目前约为5万,t 比10年前增长了10倍左右.全世界主要的苏氨酸生产企业为日本味之素公司、德国德固赛公司、美国AD M 公司、日本协合发酵工业公司等.这几大公司的产量占全球份额的90%左右.其中,日本味之素公司是世界上最大的苏氨酸生产企业,多年来该公第5期黄 金等:L-苏氨酸的生产方法及研究进展91司生产的苏氨酸占据全球市场60%以上的份额.日本协合发酵工业公司和美国AD M公司分别拥有1万t和5000t的苏氨酸生产规模.近年来,这两大企业密切合作,拟共同投资生产苏氨酸等氨基酸产品.此外,韩国希杰公司、Che il三星印尼公司等也生产苏氨酸.由于市场持续热销,各公司都有意扩大苏氨酸的产能,以满足市场需求.未来的苏氨酸市场仍有多个增长点,发展空间广阔.首先,在饲料添加剂方面的用量将持续大量增加.近10多年来,我国饲料工业产量平均年增幅达到2位数.现在全国饲料年产量已超过亿吨,成为世界第二大饲料生产国.预计到2010年,我国各种饲料添加剂的产量将达300万,t市场对苏氨酸的需求量将达到6万~8万.t其次是医药及其他领域.氨基酸大输液一直是临床用量很大的品种,苏氨酸是其主要成分之一.近年来多种氨基酸饮料正在欧美、日韩等发达国家流行,今后将向我国等发展中国家扩展.氨基酸在化妆品特别是中高档化妆品中的用量也在增多.此外,多种氨基酸片剂及口服液等保健产品现已在我国市场上崭露头角,∀瑞年#、∀万基#∀金日#等品牌的多种氨基酸保健品市场销售状况良好.各种营养食品对氨基酸的需求也在增加.参考文献:[1] 贾冬舒.苏氨酸市场现状及发展前景[J].饲料广角,2006(1):28 29.[2] 蒋莹.氨基酸的应用[M] 上海:世界图书出版公司,1999.[3] 冯美卿,瞿超进.L-苏氨酸制备方法评述[J].河北工业科技,1999,16(4):15 18.[4] Debabov V G.The threonine story[J].AdvB ioche m Eng B iotechno,l2003,79:113 136.[5] Leuchtenberger W.Am i n o acids:Techn ica lproducti o n and use[M]//Roehr M.B iotechno.l VC H,W e i n he i m,1996:465 502[6] Debabov.M ethod for preparing stra i n s w hichproduce a m i n oac i d s:U.S.A,4278765[P].1981 07 14.[7] 焦瑞身.微生物工程[M].北京:化学工业出版社,2003:16 17.[8] 常尊学,李福德.L-苏氨酸产生菌选育的研究[J].沈阳药学院学报,1990,7(3):185 188.[9] Y a m ada.Process for produc i n g L-threonineby fer m entati o n w ith Prettger.i U.S.A:5342766[P] 1994 08 30.[10]K ra m er R.Geneti c and physi o l o g ica l approaches f o r production o f a m i n o acid[J].J B i otechno,l1996,45:1 21.[11]Furuka w a S,O zaki A,Ko tan i Y,et a.lB reedi n g of L-threonine hyper producer o fE scherichia co li K12[J].B ioc h i m B i o physRes Co mmun,1988,18:788 795.[12]Ji n-H o Lee,Dong-Eun Lee,Bheong-UkLee,et a.l G lobal analyses o f transcr-ipto m es and proteom es o f a parent strain and anL-threonine-overproducing m utant stra i n[J].Jour nal of Bacteriology,2003,185:54425451.[13]Y ang-H oon K I M,Ji n-Seung PARK,Jae-Yong C HO,et a.l Proteo m ic response ana l y sis of a threon i n e-overpr oduci n g m utan tof E scherich ia co li[J].B i o che m J,2004,381:823 829.[14]Cohen G N.The co mm on pathw ay to l y si n e,m e t h ionine,and threon i n e.Am ino acids:B iosynt h esi s and genetic regu lation[M].Addison-Qesley Publish i n g Co,I nc,1983. [15]W illia m E,K arsten,Ja m es R.Purification o faspartase and aspartok i n ase-ho m oserine dehydrogenase I fro m Escherichia coli by dye-li g and chr o m atograpgy[J].Analytical B i oche m istry,1985,147:336 341.[16]M ark JM,Tho m as J S.Starvation for d ifferen tnutr i e n ts i n escher ic h ia co li resu lts i n d ifferentia lm odu lation o f rpoS levels and stab ility[J].Journa l of Bacteriology,2005,187:434 442[17]Y en-Fang K eng,Ronald E.Spec ific ity o f aspartok i n ase III fro m E scherich ia coli and anexa m i n ation of i m portant ca talytic residues[J].Archives o f B ioche m istry and B iophysics,1996,335(1):73 81.92河南工业大学学报(自然科学版)第28卷T HE METHODS AND STUDY EVOL UT I ON OFL-T HREON I NE P RODUCT I ONHUANG Ji n,XU Q i n g yang,C H E N N i n g(S chool of B ioeng i n eering,T i a njin Un i v ersit y of Science&T echnology,K ey Laboratoryo f IndustrialM icrob iology,T i a njin300222,China)Abst ract:The physicoche m ica l properties,utilization and production o f L-threon i n e w ere introduced.The producti o n m ethods,especially m icr obia l fer m entation m ethod,w ere d iscussed.Based on developm ent o f in ter national and do m estic studies,the L-threonine b i o synthesis exa m ples and m etabo lic regulation m echan is m w ere i n troduced.M oreover,the m arket perspective of L-threonine w as pr ospected.K ey w ords:L threon i n e;pr oducti o n m et h ods;study evo l u tion(上接第87页)BI OLOG I CAL RESEARCH ON CON J UGATED LI NOLE I C ACI DISO M ERASE OF M ICROORGAN IS M SWANG H ong j u n,CAO Ji a n,WANG Yu j u n,DONG L,i X I A NG L i x i n,Z HANG Lin (Colle g e of B iologicalE ngineering,H enan University of T echnology,Zhengzhou450052,Ch i n a)Abst ract:Am ong all o f the CLA iso m ers,c9,t11-18∃2and t10,c12-18∃2iso m ersw ere consi d ered to be t h e t w o w ith m any i m portant physiological activities,such as anticarci n ogen ic activity,antiather ogen i c acti v i ty,as we ll as the ab ility to reduce body-w eigh,t enhance g r ow th pro m oti o n and reduce the catabo lic effects o f i m m une sti m ulation,and thus sho w ed a w ide fie l d of applicati o n i n m edic i n e,food,healt h care and cos m etic i n dustry.So m e m i c roor gan is m s could produce con j u gated li n o le i c acid i s o m erase(EC5.2.1.5),wh ich cou l d catalyze t h e c he m ica l reaction o f li n o le ic ac i d(LA)to e ither or both of the t w o CLA iso m ers w ith the above physi o log ica l activ ities.Therefore,the production o f con j u gated li n ole ic ac i d through b iolog ical m ethod w as considered to be the develop i n g trend i n the f u ture,so con j u gated lino leic ac i d iso m erase has been attracting t h e attention o fm any researchers in and abroad.H o w ever,distribution of th is enzy m e i n species,the ir evo lu ti o nary relati o nship,its biological functi o n i n v ivo,acti v e cen ter and catalytic m echan is m etc haven%t been re vealed by now,effective expressi n g o f reco mb i n ed conjugated li n o l e ic ac i d iso m erase using genetic eng i n eering technique also re m ains to be a prob le m.Progress achieved i n the field o fb iolog ical study of con j u ga ted li n o leic acid iso m eras i n recent years,such as distribution o f conjugated li n ole ic acid iso m erase,bio l o g ica l function, proposed cata l y tic m echanis m,clon i n g and expression o f the gene w ere summ erized in t h is paper,pri m ary a nalysis o f the kno wn sequences of the enzy m e using b i o infor m atics m ethod were also put for w ard.K eywords:Con j u gated lino leic aci d iso m erase;bio l o g ical function;gene clon i n g and expressi n g;sequence a nalysis。
发酵工艺过程控制

发酵工艺过程控制发酵工艺过程控制是指在发酵过程中,通过合理控制发酵的条件和参数,使发酵过程达到预期的目标,达到最佳的产品质量和产量。
发酵工艺过程控制包括发酵条件的选择、发酵参数的控制和数据采集、分析等内容。
下面将详细介绍发酵工艺过程控制的一些关键点。
首先,在发酵工艺过程控制中,发酵条件的选择非常重要。
发酵条件包括温度、pH值、氧气供应和搅拌速度等。
不同的微生物和发酵产物有其适宜的发酵条件。
一般来说,温度是一个非常关键的参数。
适宜的温度可以提高微生物的生长速率和代谢活性,从而提高产物的产量和质量。
同时,pH值也会影响微生物的生长和代谢。
合适的pH值可以提供一个适宜的酶活性和物质转运环境。
氧气供应和搅拌速度对于需要氧气的微生物来说非常重要。
合适的氧气供应和搅拌速度可以提高微生物的通气效果和溶氧量,从而提高产量和代谢产物的选择性。
其次,在发酵工艺过程控制中,对发酵参数的控制和数据采集、分析也是至关重要的。
发酵参数包括微生物的接种量、培养基成分比例、培养基的初始pH值等。
合理的微生物接种量和培养基成分比例可以提高产物的产量和质量。
同时,初始pH值的控制可以提供一个合适的环境供微生物生长和代谢。
在发酵过程中,需要对各个参数进行实时的数据采集和分析,以便及时调整和优化发酵条件。
通过数据采集和分析,可以有效地监测发酵过程的进展,及时发现并解决问题,从而提高发酵工艺的稳定性和可控性。
另外,在发酵工艺过程控制中,发酵的时间也是需要考虑的一个因素。
发酵的时间过短可能导致微生物的生长和代谢不完全,从而影响产物的产量和质量;发酵的时间过长则可能导致微生物的过度生长和细胞死亡,影响发酵过程的稳定性和可控性。
因此,需要通过实验和经验来确定合适的发酵时间,以达到最佳的产物质量和产量。
综上所述,发酵工艺过程控制是一个非常重要的领域。
通过合理选择发酵条件和控制发酵参数,以及进行数据采集和分析,可以实现对发酵过程的有效控制,提高产品的产量和质量。
2019年苏氨酸发酵设计.doc

一、设计方案天然存在的L- 苏氨酸为无色或微黄色晶体,无臭、微甜,可溶于水,20℃时溶解度为9g/100mL,难溶于乙醇、乙醚、氯仿等有机溶剂,熔点为253~257℃。
L- 苏氨酸的解离常数为pKCOOH=2.15,pKNH2=9.12,等电点pI(25℃)=5.64。
目前,L- 苏氨酸的制备方法主要有生物合成法、化学合成法和蛋白质水解法三种。
然而,在工业化生产中,化学合成法和蛋白质水解法由于存在一些缺陷已经基本不被使用。
生物合成法则因生产成本低、资源节约、环境污染小等优点逐渐成为工业化生产L- 苏氨酸的主要方式。
生物合成法包括直接发酵法和酶转化法两种。
微生物发酵法生产苏氨酸是目前生产苏氨酸的主要方法。
采用基因工程菌进行发酵法生产,产酸可达100g/L 以上。
目前国内外已经利用微生物发酵法批量生产苏氨酸。
1.1 设计条件(1)650L苏氨酸发酵罐,分批发酵;(2)主发酵罐的尺寸及附件的设计;1.2 发酵工艺发酵法生产L-苏氨酸,通常采用短杆菌属细菌的α-氨基-β-羟基戊酸(AHV)和S-(2-氨基乙基)-L -半胱氨酸(AEC )双重抗性变异。
图一 苏氨酸发酵工艺流程1.3. 发酵罐尺寸及整体设计罐中的培养液因通气和搅拌会引起液面上升和产生泡沫,因此罐中实际装料量V 不能过大,一般取装料系数为0.6~0.75。
取装料系数o η为0.6 ,则发酵罐需装料体积为:L V V 3906.065000=⨯=⨯=η发酵罐尺寸确定发酵罐体部分的尺寸有一定的比例,罐的高度与直径之比一般为1.7~4倍左右。
1.3.1 确定发酵罐直径和高度标准式发酵罐的筒体高度和直径比:H/D 约为1.7~4 发酵罐的容量一般指圆筒体的体积加椭圆形底的体积。
V 0 = V C +2 V b Vc=(π/4) D 2 H 0V b =(π/4)D 2(h b +2/3h a )≈1.5D 3 式中:V 0—发酵罐全容量,m 3;V C —圆柱部分体积,m 3; V b —椭圆底体积,m 3; H 0—圆柱部分高度,m.因此: V 0 = V C + V b = (π/4) D 2[H 0+2(h b +1/6D)] 取 H 0/D = 2V 0 = (7π/12)D 3 +(2π/4)D 2h b可知:D=0.69m , 椭圆短半轴长度:h a =0.25D=0.25×0.69=0.1725 可知:D=0.69m则有:H 0=2D=1.38 m D i =1/3D=0.23 m S=3D i =3×0.23=0.69m C=D i =0.23 m B=0.1D=0.1×0.69=0.069 m h a =0.25D=0.25×0.69=0.173 m不同设备的厚度不同,h b 可取30 mm 、40 mm 、50 mm 。
发酵工艺对酸碱度和酒精度的调控

发酵工艺对酸碱度和酒精度的调控发酵是一种利用微生物代谢产生的酸碱度和酒精度的调控工艺。
它是一种常见的食品加工过程,可以通过调控发酵条件和微生物的数量和种类来控制产物的酸碱度和酒精度。
在发酵工艺中,微生物起着关键的作用。
不同的微生物会产生不同的代谢产物,从而影响酸碱度和酒精度。
例如,酵母菌是一种常见的发酵微生物,它可以将葡萄糖转化为乙醇和二氧化碳。
乳酸菌则会将葡萄糖转化为乳酸。
由于这些微生物的不同代谢途径和效率不同,所以产生的酒精度和酸碱度也会有所不同。
调控发酵过程中的酸碱度和酒精度通常需要考虑以下几个因素:1. 发酵条件:包括温度、pH值、氧气和营养物质等。
温度和pH值是发酵过程中最重要的因素之一,不同的微生物有不同的适宜生长温度和pH范围。
通过调整温度和pH值,可以控制微生物的生长速率和代谢产物的种类和数量,从而影响酸碱度和酒精度。
2. 微生物的选择:不同的微生物对发酵过程中的酸碱度和酒精度有不同的影响。
选择适合的微生物可以实现对产物的精确控制。
例如,如果希望获得较高酒精度的发酵产物,可以选择具有高酒精代谢能力的酵母菌。
3. 发酵时间:发酵时间对于产物的酸碱度和酒精度也有影响。
通常情况下,发酵时间越长,酒精度越高,酸碱度也会有所增加。
然而,过长的发酵时间可能会引起产物的变质和异味,所以需要根据具体情况进行调整。
通过调控发酵工艺中的这些因素,可以实现对酸碱度和酒精度的精确调控。
但是需要注意的是,发酵过程是一个复杂的生物反应过程,存在许多未知因素的干扰,所以需要根据实际情况进行调整。
此外,发酵工艺还涉及到其他一些因素,如搅拌速度、气体流速等,这些因素也会对酸碱度和酒精度的调控产生影响。
总之,发酵工艺是一种调控酸碱度和酒精度的有效方法。
通过调整发酵条件和微生物的选择,可以实现对产物的精确控制。
然而,酸碱度和酒精度的调控也需要综合考虑其他因素,并根据实际情况进行调整。
只有合理运用发酵工艺,才能保证产物的质量和安全。
第16讲 发酵过程酸碱度的控制
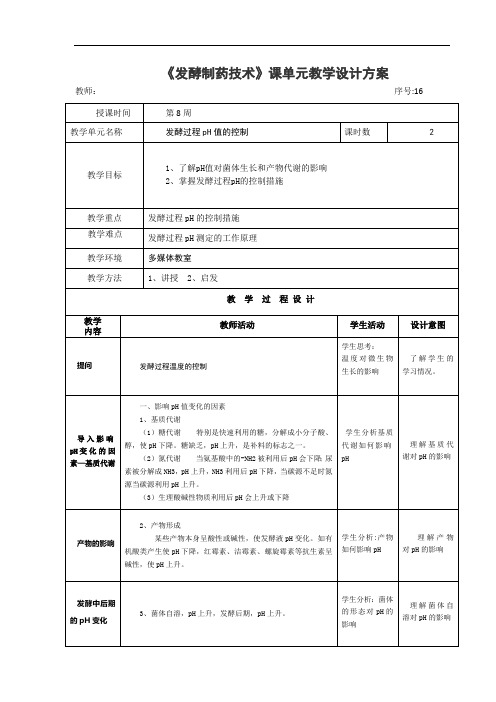
发酵过程 pH 控制
三、发酵过程 pH 值的控制 1、调整培养基的组分
学生分析培养 基的哪些成分 影响 pH
掌握发酵过 程 pH 的控制措 施
pH 的控制
2、在发酵过程中进行控制 ①添加 CaCO3: ②氨水流加法 ③尿素程 pH 的控制措 施
补料
3、通过补料调 pH
直接加酸碱 调节
课堂总结
4、当补料与调 pH 发生矛盾时,加酸碱调 pH
pH 对菌体生长和产物代谢的影响 影响 pH 的主要因素 发酵过程的 pH 控制措施
分析:补料的目 的
掌握发酵过 程 pH 的控制措 施
掌握发酵过 程 pH 的控制措 施
学生归纳总结 本次课的重点 内容
自主学习能 力的培养
教 学 过 程设计
教师活动
提问
发酵过程温度的控制
学生活动
设计意图
学生思考: 温度对微生物 生长的影响
了解学生的 学习情况。
导入影响 pH 变 化 的 因 素—基质代谢
一、影响 pH 值变化的因素 1、基质代谢 (1)糖代谢 特别是快速利用的糖,分解成小分子酸、 醇,使 pH 下降。糖缺乏,pH 上升,是补料的标志之一。 (2)氮代谢 当氨基酸中的-NH2 被利用后 pH 会下降;尿 素被分解成 NH3,pH 上升,NH3 利用后 pH 下降,当碳源不足时氮 源当碳源利用 pH 上升。 (3)生理酸碱性物质利用后 pH 会上升或下降
学生分析基质 代谢如何影响 pH
理解基质代 谢对 pH 的影响
产物的影响
2、产物形成 某些产物本身呈酸性或碱性,使发酵液 pH 变化。如有
机酸类产生使 pH 下降,红霉素、洁霉素、螺旋霉素等抗生素呈 碱性,使 pH 上升。
第六章 氨基酸的代谢控制与发酵
⑩采用低糖流加法激活PC(糖浓度为4%~5%)。
3、解除代谢互锁
在乳糖发酵短杆菌中,赖氨酸的生物合成与亮氨 酸之间存在着代谢互锁。
综上所述,在天冬氨酸族氨基酸代谢途径中,末 端产物种类多,调节机制复杂,为了高效率生产 赖氨酸,可以采取顺序解除各种调节机制的诱变 育种方法,获得多重标记突变株。
但是,采用人工诱变获得多重标记组合型突变株, 是一件费时、费力、非常麻烦的工作。采用细胞 工程和基因工程新技术,将诱变所获得的优良特 性组合起来,获得高产菌株就容易得多。
①选育Ala-突变株。 ②选育抗Asp结构类似物突变株。 ③选育适宜CO2固定酶/TCA循环酶活性比突变株。
四、蛋氨酸发酵
1、蛋氨酸高产菌应具备的生化特征
⑴CO2固定反应能力强。
⑵Asp合成能力强。 ⑶AK活力强。
⑷HD活力强。
⑸PS活力微弱或丧失。 ⑹高丝氨酸激酶活力微弱或丧失。 ⑺GHD活力弱。 ⑻O-琥珀酰高丝氨酸转琥珀酰酶活力强。
⑴切断支路代谢
①选育Lys-、Lysl、Lys+突变株。
②选育Thr-、Thrl、Thr+突变株。
⑵解除反馈调节
①选育AECr和AHVr突变株。
②选育抗SAM结构类似物突变株。
③选育抗蛋氨酸结构类似物(如乙硫氨酸、硒代 蛋氨酸、1,2,4-三唑、三氟蛋氨酸)突变株。
⑶切断蛋氨酸向下反应的通路
选育SAM-突变株。
丙氨酸是比较活跃的氨基酸,在生物体内通过转 氨作用可生成其它氨基酸,生成丙氨酸的途径必 然消耗许多PEP。
发酵过程控制氨基酸发酵的工艺控制实例
(二)温度的影响及其控制
1、温度对Glu发酵的影响
谷氨酸发酵前期(0~12h)是菌体大量繁殖阶段,在此 阶段菌体利用培养基中的营养物质来合成核酸、蛋白质等, 供菌体繁殖用,而控制这些合成反应的最适温度均在 30~32℃。
在发酵中、后期,是谷氨酸大量积累的阶段,而催化 谷氨酸合成的谷氨酸脱氢酶的最适温度在32~36℃,故发 酵中、后期适当提高罐温对积累谷氨酸有利。
在发酵后期由于耗用大量NH4+ ,pH值下降, 此 时就要进行pH值调节,以保证发酵的正常进行。
pH发生变化的主要原因是培养基中营养成分的 利用和代谢产物的积累。
如当谷氨酸棒状杆菌利用糖类物质不断生成谷氨酸时,培养 液的pH就会下降。而酸性物质的消耗和氨的生成等则会导致培 养液的pH上升。
pH:前期pH7.5~8.0,中后期pH7.0~7.6。 通过采用流加尿素,氨水或液氨等办法调节 pH,补充氮源。
表1 部分氨基酸及其生产所用菌株
氨基酸 谷氨酸
缬氨酸 丙氨酸 脯氨酸 赖氨酸
苏氨酸 鸟氨酸 亮氨酸 酪氨酸
使用的菌株
谷氨酸棒杆菌、乳糖发酵短杆菌或黄色短 菌杆、北京棒杆菌、钝齿棒杆菌 北京棒杆菌、乳糖发酵短杆菌 凝结芽孢杆菌 链形寇氏杆菌 、黄色短杆菌 黄色短杆菌 、乳糖发醇短杆菌 、谷氨酸棒 杆菌 大肠杆菌、大肠杆菌 谷氨酸棒杆菌 、黄色短杆菌 黄色短杆菌 氨酸棒杆菌
二次世界大战后不久,美国有人提出用 发酵法生产谷氨酸的报告。
日本也相继开始了研究,1956 年日本 协和发酵公司分离出一种新的细菌,它可 以利用100 克葡萄糖转化为40 克以上的谷 氨酸。1957 年发酵法味精正式商业性生产, 这标志着氨基酸发酵工业的诞生。
生产氨基酸的大国为日本和德国。
L-苏氨酸的生产工艺
株 。其 对菌 种 的生 产能 力和 发酵设 备及 发酵条 件要
求 都较 高 ,因而生 产往 往受 到限制 。
22 化 学合成 法 .
苏 氨酸 的化学 合成路 线根 据原料 的不 同可 分为
巴豆酸法 、乙酰 乙酸 乙酯 法 、不 对称 环氧 化法 及甘
天 然存 在 的 L 苏 氨 酸 为 无 色 或 微 黄 色 晶 体 , 一
L 一苏 氨 酸 的 生 产 工 艺
冯 烁
(南京 农 业 大 学 动物 医学 院 ,南 京 209 ) 10 5
摘
要 :文 章 介 绍 了 苏氨 酸 的理 化 性 质 ,重 点介 绍 了 苏氨 酸 的 不 同生 产 3 艺特 点 、 工 艺 流程 ,影 响 生 产 工 艺 -
的 因素 ,并 展 望 了苏氨 酸 的 市场 前 景 。
无臭 、微 甜 ,可溶 于水 ,难溶 于 乙醇 、乙醚 、氯仿 等 有 机 溶 剂 ,2 【 溶 解度 为 9g 10m ~ 0c =时 ・0 L ,熔
的 8种必需 氨基 酸 中 ,苏氨 酸仅 次于蛋 氨酸 、赖 氨 酸 和色氨 酸 。在 畜禽饲料 中添加苏 氨酸 能提 高饲料 蛋 白质 的生 物学 价值 ,平 衡各 种氨基 酸 ,促进 蛋 白
道 并不 多 。直 接 发 酵法 生 产 L 苏 氨酸 ,通 常采 用 一 短 杆 菌属 细 菌 的 一 基 一 氨 羟基 戊 酸 ( V) S AH 和 一
( - 基 乙 基 ) L 半 胱 氨 酸 ( E 双 重 抗 性 变 异 2氨 一一 A C)
苏 氨 酸 的 分 子 式 为 CHN 。O ,结 构 式 为 C , H一
反应 条 件缓 和 、产率 高 ,缺 点是 步骤 繁琐 、溶 剂成
代谢控制发酵-第六章 代谢控制发酵实例
(2) 选育渗漏缺陷型突变株 苯丙氨酸和酪氨酸双营养缺陷(或预苯酸缺陷)使得菌株生长
缓慢,因此可选育预苯酸渗漏缺陷型突变株 渗漏缺陷型是一种不完全营养缺陷型,它不会产生过量的末
端产物,因而可以避开反馈调节 但它又能合成微量的末端产物,用来进行生物合成 在培养这种突变体时,可不必在培养基中添加相应的物质,
弱化子
p 解除反馈调节,增强色氨酸合成途径代谢流(通) 解除色氨酸抑制作用:
(1) 选育色氨酸结构类似物(5-氟色氨酸、5-甲基色氨酸) 抗性突变株
变构,失活
酶
色氨酸
问题:诱变后存活的微生物 包括正向突变(想要的)、负 向突变(不想要的),怎么把 正向突变的筛选出来?
5-氟色氨酸 5-甲基色氨酸
四、青霉素发酵
生物合成途径
四、青霉素发酵
代谢调节
碳分解代谢产物的影响
Ø 青霉素的生物合成受碳分解代谢产物阻遏,如ACV合成 酶,IPN合成酶,酰基转移酶就被阻遏
Ø 葡萄糖可以刺激菌体生长,使作为赖氨酸和青霉素合 成中间体的α-氨基己二酸转向合成赖氨酸,抑制青霉 素的合成
Ø 葡萄糖降低青霉素生物合成的速率和得率还由于葡萄 糖与6-APA之间形成复合物,从而减少了可用于合成青 霉素的中间产物。
Ø 选育单氟乙酸、三氟乙酸敏感突变株 抑制乌头酸梅和异柠檬酸脱氢酶活性。若菌体对药品 敏感,说明该突变株的乌头酸酶和异柠檬酸酶活力低或 含量少
Ø 选育强化CO2固定反应的突变株 氟丙酮酸敏感、天冬氨酸缺陷、羧化酶基因克隆
Ø 强化柠檬合成酶
一、柠檬酸发酵
发酵控制
Ø 控制Mn2+和NH4+浓度,解除柠檬酸对PFK的抑制,使EMP 畅通无阻