水稻黄绿叶基因的克隆及应用
211055772_水稻抗稻瘟病免疫机制分析与抗病育种措施应用

粮油农资 162023.3持续发展。
首先,要注重宣传稻田有机肥施用技术,建议根据当地土壤性质,采用适当的比例,开展化肥减量替代。
其次,绿色种养循环是有效推进水稻化肥减量增效、积极应对施肥成本上升的迫切需要,更是推进控污减排、促进水稻绿色低碳发展的迫切需要。
因此,推广水稻化肥减量技术,降低施肥强度和施肥次数,可达到稻田化肥减量增产的效果,助力江苏沿海地区水稻绿色高效可持续生产。
作者简介:赵燕芬(1973-),女,本科,农艺师。
研究方向:作物栽培技术推广和服务。
水稻抗稻瘟病免疫机制分析与抗病育种措施应用王春红(吉林省德惠市大房身镇综合服务中心,吉林 德惠 130300)根据国家统计局数据显示,我国水稻产出大省是黑龙江、湖南和江西,均已超过2000万吨。
黑龙江2020年全省水稻产量为2896.15万吨,2021年水稻产量则达到2913.7万吨,同比增长了0.61%。
而湖南水稻产量呈现明显波动增长趋势,2020年水稻产量2638.94万吨,而2021年产量为2683.1万吨,同比增长为1.67%。
江西水稻产量整体呈现先上升后下降,2020年产量为2051.2万吨,2021年则为2073.9万吨。
在水稻作物种植过程中,常会出现较为严重的病虫害,其中最为严重的典型水稻病害为抗稻瘟病,大幅度降低水稻产量,需要重点研究水稻抗稻瘟病免疫机制,并制定出科学有效的抗病育种策略。
1 水稻抗稻瘟病免疫机制分析以黑龙江省水稻种植为例,黑龙江省水稻种植面积超过400万公顷,稻瘟病发病严重。
其中,穗颈瘟发病比例为9%~10.3%,区域比例种植达到65%以上。
该品种苗瘟9级、叶瘟7级、穗颈瘟9级,经过近年来综合抗病品种研究得出,长期种植单一品种,田间会存有大量对该品种致病性较强的菌源,促使侵染该品种的生理小种变为优势小种或稻瘟病菌生理小种产生变异。
稻瘟病可以发生在水稻生长的任何一段时期内,而且其发病部位较多,包括苗、节、叶和稻穗等诸多部分,会严重威胁水稻生长质量和产量。
水稻抗白叶枯病基因的挖掘及其功能研究进展

5 0 %的产量 损失 。它 于 1 9世 纪 9 0年代 初 .首 次 在 日本 地 区发 现 .随后在 世界其 它各 主要 稻区 相继 被报 道 ,如 非洲 、亚洲 以及澳 洲等地 。中 国早 在上
( X a n t h o m o n a ¥ o r y z a e p v . o r y z o e 。 X o o )引 起 的水 稻
各地却 又相 继爆 发 白叶枯病 。究 其原 因 ,很 可能 就 是 由于 xo o生 理 小 种 的 自然 突变 导致 原 有 抗 病 品
白叶枯病 ( b a c t e r i a l b l i g h t , B B)是 水稻 生 产过 程 中
一
种抗性 的丧 失 .从而 导致 白叶枯 病 的 “ 死 灰复燃 ” 。
因此 .需要 不断 的培 育新 的抗病 品种 来提 升水 稻 的
抗病 能力
种 重要 的病 害 。尽 管 目前 已经就这 种病 害的 防治
和抗 性品种 的培育 做 了大量 的工作 .但病 害依 旧会
悄 悄 卷土重来 。从 长期 的实践来 看 .培育新 的抗病 品种 和挖掘 新 的抗 病相关 基 因仍是控 制这些 病 害的
Xa 2 1基 因编 码 一 个植 物 类 受 体蛋 白激 酶 .此 前 的 研究 表 明该酶在 多 种生物 系统 中介 导细 胞 内的信号 传 导 .而 信号传 导 被认为 是植物抵 抗病 毒 、细菌 以
片处 的病 斑 会 逐 渐 变枯 变 长 .且 由绿 色 变 为 黄 色
[ 3 1
。
及 真菌 的一个 重 要 机制 该 激 酶 由 1 0 2 5个 氨 基 酸
基于KASP技术用于水稻产量基因分型的引物组合及其应用[发明专利]
![基于KASP技术用于水稻产量基因分型的引物组合及其应用[发明专利]](https://img.taocdn.com/s3/m/98df3fdc5ff7ba0d4a7302768e9951e79b8969d1.png)
(19)中华人民共和国国家知识产权局(12)发明专利申请(10)申请公布号 (43)申请公布日 (21)申请号 201710297282.X(22)申请日 2017.04.28(71)申请人 华中农业大学地址 430070 湖北省武汉市洪山区狮子山街特1号华中农业大学(72)发明人 余四斌 袁志阳 孙文强 熊银 龚蓉 张佳宁 (74)专利代理机构 北京路浩知识产权代理有限公司 11002代理人 王文君(51)Int.Cl.C12Q 1/68(2006.01)C12N 15/11(2006.01)(54)发明名称基于KASP技术用于水稻产量基因分型的引物组合及其应用(57)摘要本发明提供一套基于KASP技术开发的用于水稻产量基因分型的引物组合,所述水稻产量基因包括Gn1a、NAL1、Ghd7、Hd1、Ghd7.1和Ghd8,所述引物组合的序列分别如SEQ ID NO:1-18所示。
利用本发明提供的成套KASP功能标记及其引物组合能够快速检测水稻种质中产量基因功能等位基因的组合方式,检测结果准确可靠,操作简单,成本低。
本发明方法能够实现水稻育种材料重要产量功能基因的检测,为水稻高产新品种选育、改良提供科学指导。
权利要求书2页 说明书8页序列表3页 附图4页CN 106939349 A 2017.07.11C N 106939349A1.基于KASP技术开发的用于水稻产量基因分型的引物组合,其特征在于,所述水稻产量基因包括Gn1a、NAL1、Ghd7、Hd1、Ghd7.1和Ghd8,它们的功能性分子标记分别对应于表1中M1~M6,信息如下:表1编分子标记类型物理位置等位基因号M1SNP LOC_Os01g10110基因编码区第1605位[T/G]M2SNP LOC_Os04g52479基因编码区第698位[A/G]M3Indels LOC_Os06g1637基因编码区第1053-1056位[AAAG/-]M4SNP LOC_Os07g15770基因编码区第455位[A/G]M5Indels LOC_Os07g49460基因编码区第1516-1523位[AACGTTGT/-] M6SNP LOC_Os08g07740基因编码区第566位[A/C]基于KASP技术的用于检测上述功能性分子标记的引物组合,所述引物组合的信息见表2:表22.含有权利要求1所述引物组合的用于水稻产量基因分型的检测试剂盒。
水稻耐盐碱性生理和遗传研究进展

水稻耐盐碱性生理和遗传研究进展植物遗传资源2007,8(4):486~493JournalofPlantGeneticResources水稻耐盐碱性生理和遗传研究进展祁栋灵,郭桂珍,李明哲,曹桂兰,张俊国,周庆阳,张三元,徐锡哲,韩龙植('中国农业科学院作物科学研究所/国家农作物基因资源与基因改良重大科学工程/农业部作物种质资源与生物技术重点开放实验室,北京100081;.中国热带农业科学院橡胶研究所,海南儋州571737;3吉林省农业科学院水稻研究所,公主岭136100;韩国农村振兴厅农业生命工学研究院,韩国水原441~707;四川农业大学,雅安625014)摘要:水稻是对盐碱中度敏感的作物,其耐盐碱性因品种,生育阶段,器官,土壤盐碱类型等而存在差异.盐碱胁迫对水稻的伤害主要表现为延迟种子发芽时间,降低发芽率,抑制生育进程,阻碍幼穗分化,推迟分蘖时间,减少分蘖数,降低产量和品质.本文论述了近年来在水稻耐盐碱生理机理,转运蛋白,遗传和数量性状位点的分子检测,分子信号传导以及基因克隆转化方面的研究进展,并对今后的研究工作提出了建议.关键词:水稻;耐盐碱性;生理;遗传;数量性状位点ProgressofPhysiologySaline.AlkalineandGeneticResearchonToleranceinRiceQIDong.1ing,,GUOGui.zhen,LEEMyung-chul,CAOGui.1an,ZHANGJunguo, ZHOUQing—yang,ZHANGSan—yuan,SUHSeok-cheol,HANLong—zhi (KeyLaboratoryofCropGermplasmResourcesandBiotechnology,MinistryofAgriculture ,/eNationalKeyFacility-厂0rCropGeneResourcesandGenet~Improvement~InstituteofCropScience,ChineseAcade myofAgriculturalSciences,eng100081;InstituteofRubberResearch,ChineseAcademyTropicalofAgriculturalScienc es,Danzhou571737;InstituteofRiceResearch.JilinAcademyofAgriculturalSciences,Gongzhuling136100;Na tionalInstituteofAgriculturalBiotechnology,RDA,Suwon441—707,Korea;SichuanAgriculturalUniversity,Y aan625014)Abstract:RiceiSoneofthemoderatesaline-alkalinesensitivenesscrops.Thedifferentkinds Ofvarieties,growthstage,organandsoilofsaline—alkalineresultinthedissimilaritiesofsaline—alkalinetolerance.Whenricesufferedfromsaline—alkalinestress,seedsproutingpostponed,seedingratedecreased,growthanddevelopmentin —hibited,paniclesinitiationhindered,tilleringdelayed,totaltilleringnumbers,yieldsandqual itydecreased.Therecentprogressesofphysiologicalmechanism,transporter,genetic,QTLanalysisbasedonm olecularmarkers,mo—lecularsignalconduction,genecloneandtransformforsaline—alkalinetoleranceinricewerereviewed,andthere—searchdirectioninthefuturewasdiscussed.Keywords:Rice;Saline—alkalinetolerance;Physiology;Genetic;QTL盐碱危害是影响全球大田作物生产的严重问题,它妨碍作物的正常生长,限制作物产量潜力的正常发挥.据统计,在影响作物产量的各种环境因素中,干旱和盐碱所造成的减产程度高达40%以上.全球约有9×10.hm的土地存在不同程度的盐碱化,占世界总耕地面积的20%Ea3,中国盐碱土约为2000万hm[23.随着中国人口的不断增加和城镇化的迅速发展,中国可耕地面积不断减少;干旱的频收稿日期:2006.9—28修回日期:2007—08—24基金项目:国家科技支撑项目(2006BAD13BO1);农业行业标准项目(05177);作物种质资源保护项目[NB06—070401(22—27)一O1];中韩合作项目(2007~2009)作者简介:祁栋灵,河南太康县人,硕士,主要从事水稻耐盐碱性遗传研究.通讯作者:韩龙植,博士,研究员,博导.Tel:010—62176784,E—mail:********************4期祁栋灵等:水稻耐盐碱性生理和遗传研究进展487繁发生和灌溉方法的不当造成了次生盐碱化面积日益扩大,再加上近年淡水资源严重不足,中国干旱,半干旱及滨海地区的粮食生产受到严重威胁.因此,如何开发和利用盐碱化土壤,提高作物产量和品质已成为盐碱地农业,海水灌溉农业中急待解决的重要问题.水稻是一种对盐碱中度敏感的作物,土壤盐碱化是盐碱稻作区水稻生产稳定发展的主要限制因素.水稻耐盐碱性是指在盐碱环境下水稻忍耐或抵抗盐碱胁迫的能力.深入开展水稻耐盐碱性研究,对发挥水稻品种在盐碱稻作区的产量潜力,进一步扩大水稻的可种植面积,保证盐碱稻作区粮食的安全生产和提高人民的生活水平以及改善生态环境具有十分重要的意义.本文论述了水稻耐盐碱性的遗传差异,盐碱胁迫对水稻生长发育和农艺性状的影响,水稻耐盐碱生理机理,遗传和数量性状位点的分子检测,分子信号传导以及基因克隆转化等方面近年来的研究进展,以期为进一步深入开展水稻耐盐碱性研究提供参考.1水稻不同基因型,不同器官和不同发育时期耐盐碱性差异1.1水稻耐盐碱能力水稻起源于淡水沼泽地区,属于对盐碱胁迫中度敏感的一类作物J.试验表明,水稻种子在萌发期间用200mmol/L的NaCI溶液处理,明显抑制幼苗的根长,芽长和根数;当水稻生长到4叶期时,用283mmol/L的碳酸钠溶液进行胁迫处理,稻苗迅速大量死亡,189mmol/L碳酸钠溶液可作为水稻苗期耐盐碱性鉴定浓度.梁正伟等研究表明,在pH9.49的苏打盐碱土中水稻单株分蘖力明显下降; 程广有等指出,pH8.6为水稻品种分蘖盛期和孕穗期耐碱鉴定的临界pH值,可作为这两个时期水稻耐碱性鉴定的胁迫条件.同一水稻品种的耐盐性与耐碱性存在一定的差异.程广有等利用模拟盐碱池对水稻品种在分蘖盛期和孕穗期进行耐盐性与耐碱性相关性研究表明,水稻品种的耐盐性和耐碱性问存在一定差异,即耐盐性强的水稻品种未必表现为较强的耐碱性,反之亦然.1.2水稻品种间耐盐碱性差异对13029份水稻种质进行苗期耐盐性鉴定结果表明,耐盐性呈严重的偏态分布,水稻种质间存在较大的耐盐性差异;籼稻品种中的耐盐种质数量多于粳稻,粘稻品种中的耐盐种质数量多于糯稻,水稻品种中的耐盐种质数量多于陆稻.张秀双等¨研究表明,不同粳稻品种之间的耐盐限度存在差异.水稻种质问的耐盐性差异,可能与各自的历史起源,演化和遗传背景等因素有着密切的关系.1.3水稻不同发育时期耐盐碱性差异郭望模等¨研究表明,水稻在种子萌发期表现为相对耐盐,但盐浓度过高会延迟种子萌发,幼苗期对盐碱较敏感,同一品种在发芽期和幼苗期的耐盐性存在一定差异.Kbar121指出,水稻植株进入营养生长阶段,对盐碱的耐性逐渐增强,而到生殖生长期,对盐碱胁迫又变得敏感.不同生育阶段耐盐碱性的差异,可能与水稻自身遗传调控,生理生化特点密切相关.1.4水稻植株器官间耐盐碱性差异韩朝红等研究表明,水稻成熟期的株高和秆长对盐胁迫反应较迟钝,而发育旺盛的器官如新叶,幼根和花穗等器官对盐较敏感,水稻幼苗地上部较地下部对盐胁迫反应敏感.梁正伟等研究指出,在盐碱胁迫下,水稻成熟期的株高和秆长不能很好地反映品种间耐盐碱性强弱,而单株分蘖数是衡量水稻耐盐碱强弱的良好指标.王建飞等¨研究认为,在相同温度和盐胁迫浓度下,幼苗期的叶片盐害级别,相对苗高和相对茎叶干重等在不同水稻品种间存在显着差异.2盐碱胁迫对水稻生长发育及农艺性状的影响在不同土壤和水环境条件下,定量监测结果表明,水稻植株体中高背景值的Na,HCO一及重碳酸根离子会严重影响水稻对土壤中营养成分的吸收.水稻体内钠离子过量积累,将对微量元素和营养元素的利用产生强烈的拮抗作用,进而导致作物根部和地上部顶端分生组织的生长受阻,并影响细胞壁多糖的生物合成,从而影响细胞正常分裂和细胞伸长.同时,对叶绿素的合成产生影响,易导致作物叶片变成不正常的暗绿色,引起作物生理失调,正常代谢受阻,造成作物生长弱小,结实率降低,呼吸强度减弱,使正常生长发育受到阻碍,最终影响水稻农艺性状的表型值.2.1萌芽与出苗Girdhar¨研究表明,盐分可延迟水稻种子的萌芽和出苗.佟立纯等…报道,水稻种子在萌发阶段受到盐碱胁迫,常会导致种子发芽不齐,发芽势降低,发芽率下降.但到目前为止,盐胁迫对种子萌发488植物遗传资源8卷的作用机理仍不十分清楚.秦忠彬等¨引指出,盐分对水稻萌发期的影响主要是限制种子的生理吸水. 闫先菩等¨则认为,在种子吸胀过程中盐胁迫会破坏细胞膜,使其透性增大,导致溶质外渗,种子萌发受阻.在种子萌发和出苗过程中,体内储存的有机物质被分解和转化,新的有机物又被合成.NaCI胁迫可能会影响这些代谢过程中的酶活性,特别是脂肪分解过程中的脂肪酶等,进而干扰或破坏这些酶所参与的正常的生理代谢过程.2.2幼苗生长出苗以后,水稻从异养阶段向自养阶段过渡.此时如果遇到盐碱胁迫,则会引起叶片卷缩和枯萎, 叶尖变黄,并抑制叶片的伸长,新生叶的形成和新根发出,从而导致侧根和根长的显着减少以及苗高的降低.浓度高时,会导致幼苗死亡口.'.2.3生长发育和成熟光合作用是形成光合植物生物量的前提,是生长发育的物质和能量基础.所有影响光合作用的因素都会间接影响植物生物量的积累和经济产量.盐碱胁迫是影响植物光合作用的主要不利因素之一.在盐碱胁迫下,分蘖高峰明显推迟或不出现分蘖高峰,抽穗期延长,不耐盐碱的早熟品种比耐盐碱的中熟品种抽穗晚;水稻株高降低,单茎绿叶数和有效分蘖数减少.Lee等副研究表明,水稻叶片在分蘖期受害最严重,茎秆和花序的长度在孕穗期严重缩短,有效穗数,千粒重和分蘖数等产量构成指标明显减少.研究结果表明,耐盐碱性差的水稻秧苗,在光合作用时受盐碱离子胁迫的影响较大,这些离子主要是通过干扰气孔运动,减少CO:摄入量来影响光合作用.盐胁迫下,胞内CO:分压上升,不利于CO,进入细胞;膨压下降,保卫细胞正常的形态受到破坏,气孔导性减弱,cO:净同化率降低.李海波等认为气孔关闭是水稻敏盐品种在盐胁迫下光合能力下降的主要原因.盐碱胁迫通过阻碍植物的光合效率,减少光合产物的有效积累,运输和分配,最终影响生长发育和成熟期各种生理生化代谢活动的正常进行.水稻在生殖阶段受到外界盐碱胁迫,将导致水稻花粉活力的降低,从而干扰授粉和花粉粒的萌发过程.水稻盐碱敏感品种淀粉合成酶的活力容易受到抑制,幼穗分化严重受阻碍,结实率显着降低;盐碱胁迫严重影响幼穗的正常分化和小穗的形成, 导致空秕率的增加,尤其对幼穗分化的影响最大,显着缩短或减少幼穗长度,每个小穗的第一枝梗数,小穗数,着粒密度,谷粒的长度,厚度,宽度,千粒重及小穗重量,且导致稻草和谷粒产量的下降以及稻米品质的降低.3水稻耐盐碱生理机理盐碱环境对植物产生两种胁迫,即渗透胁迫和离子胁迫.植物若要在盐碱胁迫的环境中正常生长,就必须具有克服这两种胁迫的能力...作物的耐盐碱能力取决于自身的遗传特性及其所在的环境条件.高等植物的耐盐碱性包含聚集盐碱离子和排除盐碱离子两个方面.一般非盐生植物多属于后者,而限制盐碱离子进入地上部是决定其耐盐碱能力强弱的一个重要方面.高浓度的盐碱离子通过根际环境借助根系的吸收,转运蛋白运载和跨膜运输等环节最终影响作物的生长发育过程.3.1无机离子吸收运输调节郑少玲等研究表明,与敏盐水稻品种相比,耐盐水稻品种的中柱薄壁细胞对Na有相对积累作用,从而控制Na向地上部的运转,减少对地上部组织的盐害,进而提高自身耐盐性.Baba等对水稻24h短期盐胁迫处理结果表明,水稻木质部内的溶液离子浓度,如K,ca",Mg"的浓度显着提高,体内的Na转运被抑制,K数量迅速增加,表现出较高的离子调节能力.Hussan等用不同浓度的氯化钠溶液处理水稻幼苗结果表明,根干重显着减少,钾离子浓度在地上部组织中显着降低,在茎和根中的Na/K比率下降,认为离子选择性吸收是主要的耐盐机理.Lin等在盐胁迫下对水稻根茎染色体片段研究结果表明,敏盐水稻品种的叶片盐害是从根部转运到茎部,并在茎部大量积累的Na所造成. 3.2渗透调节渗透胁迫是限制水稻及其他作物产量的主要非生物因素之一,而渗透调节能力则是植物耐盐碱的最基本特征之一.从细胞水平上讲,耐盐碱能力的强弱在很大程度上取决于细胞自身的渗透调节能力.植物受到渗透胁迫造成的不平衡,通常在细胞内积累渗透保护物质,以降低细胞的渗透势.在盐胁迫条件下水稻幼苗能通过自身细胞的渗透调节作用,如在细胞内合成脯氨酸,可溶性糖,果糖,蔗糖,多胺等具有渗透调节功能和较强亲水力的相容性溶质,以保护细胞中蛋白质,蛋白复合物和膜结构免遭降解或破坏,从而使细胞维持正常的生理活动,适应外界渗透胁迫环境,缓解盐害,进而提4期祁栋灵等:水稻耐盐碱性生理和遗传研究进展489 高水稻幼苗耐盐性….盐碱胁迫下,在水稻叶片中会累积大量的脯氨酸,脯氨酸积累的多少可反映水稻幼苗伤害程度.Nguyen等用100mmol/LNaC1溶液处理水稻幼苗6d后,发现在植株茎内盐诱导出一些含氮化合物,包括游离氨基酸和胺脲类物质,尤其脯氨酸含量显着增加,并且游离氨基酸在茎的累积量与Na浓度高度正相关.谢国生等指出,盐碱胁迫下水稻幼苗脯氨酸含量是反映水稻幼苗盐碱胁迫程度的重要指标之一.刘春玲等发现,盐胁迫下各水稻品种内源游离脱落酸含量迅速积累,耐性品种积累的脱落酸量都较敏感品种高,并且耐性品种ABA积累的时间较长.Hien等指出,脯氨酸积累是由渗透压诱导合成的结果,根中脯氨酸累积是水稻品种耐渗透胁迫的生理指标之一.在逆境条件下,植物体内积累脯氨酸具有一定普遍性,脯氨酸可能作为渗透调节物,质膜和酶的保护物质以及自由基消除剂等,对植物起保护作用.3.3转运蛋白及跨膜运输高等植物的膜系统上普遍存在HATPase,Na/H转运蛋白和水孔蛋白等膜蛋白,这些转运蛋白对细胞内离子分布和细胞水势具有调节和信号传导功能,对细胞与环境物质交换和信息交流起关键性的介导作用,而且植物体内Na,cl,H和K等离子的跨膜运输也主要由转运蛋白来完成.高等植物在逆境胁迫响应下的渗透调节过程中,膜和液泡上的H一ATPase具有维持细胞内的Na,Cl一浓度的功能,水稻Na/H转运蛋白(OsNHX1)在离子跨膜运输和细胞盐区隔化中起着关键作用.近年来,人们发现了很多与盐胁迫有关的转运蛋白,对它们的作用机理开展了初步地研究和分析. Nakamura等¨在野生稻叶绿体的类囊体膜上发现了一种特异性转运蛋白,该转运蛋白参与水稻的耐盐反应.由OsNHX1基因编码液泡膜上Na/H的逆转运蛋白,在高浓度下,可以提高水稻茎和根中的OsNHX1转录水平,OsNHX1的过量表达可提高水稻细胞和植株水平上的耐盐性.Karuna等认为,Na/H逆转运蛋白的数量是决定水稻耐盐性强弱的最重要的因素之一.在水稻盐胁迫下,晚期胚胎发生富集蛋白(LEA蛋白)作为脱水保护剂在脱水的营养组织中能够受盐胁迫诱导而迅速积累,保护细胞免遭水势降低的胁迫损伤,而在外界盐胁迫解除后LEA蛋白积累量会逐渐减少.这种在盐胁迫下诱导合成积累,在盐胁迫解除后又减少合成和积累,可能是转运蛋白的一大特征.一些学者研究表明,水稻体内的Na和K的吸收由不同的转运蛋白来完成,在K亏缺时,OSHKT1在水稻根中特异性介导Na的转运,而不转运K,TAHKT1可转运K和Na.这表明水稻体内的Na和K可能具有不同的吸收途径和转运机理.4水稻耐盐碱性遗传遗传研究表明,水稻耐盐碱性是多种耐盐碱生理反应的综合表现,是由位于不同染色体上的多个基因所控制的数量性状_5.Moeljo等认为盐胁迫下水稻单株死叶率和茎叶干重的遗传变异主要表现为等位基因间的加性效应及互作.Jones等认为水稻苗期耐盐性可能由少数几个基因控制,其遗传变异来源于加性和显性效应,并以加性遗传效应为主,没有发现上位性互作.Akbar等研究盐胁迫下水稻苗期多个性状的遗传变异结果表明,苗高, 地上部Na和Cl一含量,茎叶干重,根部干重具有较强的基因加性效应,遗传力较高;而根长和根部Na,CI一含量的遗传力较低,基因的显性作用占优势;并推测至少有3对基因控制茎叶Na,cl一含量的遗传,有2对基因控制根长,茎叶和根干重.顾兴友等利用耐盐品种Pokkali和敏盐品种Peta配制回交群体,检测到影响水稻苗期耐盐性的4个数量性状位点,其增效等位基因均来自耐盐品种,影响成熟期耐盐性的数量性状位点的有利基因来自于双亲;RG678和RZ400B~RZ792附近的2个数量性状位点在全生育期均表达出较强的耐盐性,并且指出水稻苗期和成熟期耐盐性存在共同的遗传基础.郭岩等获得耐盐性可稳定遗传到第13代的变异系,并且指出耐盐性受一个主效基因控制.以上对水稻耐盐性的遗传分析多以苗期为主,并且不同的研究者所采用的品种和耐盐指标不尽相同,但发现对盐胁迫产生反应的大多数指标均表现为数量性状特征.可以看出,水稻耐盐性的遗传变异主要由基因加性效应和显性效应控制,这两种效应是构成水稻苗期耐盐性的重要遗传基础.在水稻杂交后代耐盐性遗传方面,有些学者认为,其杂种优势耐盐性状不明显,至于原因仍不十分清楚.5水稻耐盐碱性分子机理5.1分子定位利用现代分子生物学技术和日益完善的统计方490植物遗传资源8卷法,许多学者对水稻耐盐碱数量性状进行了分子定位研究.龚继明等'利用一个加倍单倍体群体和由其构建的遗传图谱进行了水稻耐盐基因定位研究,发现耐盐性呈正态分布,属于典型的数量性状,并检测到一个主效QTL_sId和7个微效QTL,QTL. std位于第1染色体的RG612和C131之间,能解释15.6%的表型变异.Lin等在盐胁迫下检测到与幼苗存活率有关的3个QTL,分别分布在第1,6和7 染色体上,其中两个为主效QTL,即SNC.7和SKC. 1,两者可分别解释48.5%和40.1%的表型变异.顾兴友等利用耐盐品种和敏盐品种配制回交群体,检测到影响水稻苗期耐盐性的4个QTL,分别分布在第5,6,7和9染色体上.Zhang等利用表1水稻耐盐性QTL定位现状Table1StatusofQTLanalysisforsalttoleranceinrice RFLP标记在水稻第7染色体上检测到1个耐盐相关QTL.在盐胁迫条件下,林鸿宣等在第5染色体上的RG13位点检测到一个与幼苗存活天数相关的QTL,其表型贡献率达到11.6%.Prasad等叫在第6染色体上检测到7个耐盐QTL.Koyama等1] 对水稻植株茎中的Na和K吸收量,Na和K的浓度,Na和K的浓度之比等5个性状进行了QTL分析,共检测到11个与耐盐性有关的QTL.目前,在水稻12条染色体上检测到的耐盐QTL已有30多个,在水稻第6和7染色体上检测到的耐盐性QTL最多,其次是第1和第5染色体,而至今在第11染色体上未检测到与耐盐性相关的QTL (表1).耐盐性状SalttolerancetraitParentpulationandma类rk.Q.Reference幼苗存活天数SDSNa浓度NCK浓度KCNa吸收量NUK吸收量KUNa:K吸收比NKR苗干物重DM发芽率SG苗茎长SSL苗干物重SDM苗耐盐性SV苗期盐害级别RSI苗Na含量NC鲜重/干重SFW/DMW 成熟期茎叶重SW结实率SSR主穗长MPL株高PH主穗颖花数NSMP幼苗存活天数SDS苗平均存活天数ASDS 单株活力与育性耐盐级别总数ISSVF NonaBokra×Koshihikari特三矮2号×CB Tesanai2×CB窄叶青8×京系l7 ZYQ8×Jx17M一2O×77一l70 6.4l9.68.8lO.68.96.8一l9.69.19.69.7l6.3~l9.5l8.9l3.5~l7.9l6.3Lin[351Koyama[]Pr跚d[删顾兴友[林鸿宣】龚继明[57]Zhang[]SDS:Survivaldaysofseedling;NC:Naconcentration;KC:Kconcentration;NU:Nauptake; KU:Kuptake;NKR:Na:Kratio;DM:Drymass;SG:Seedgermination;SSL:Seedlingshootlength;SDM:Seedlingdrymatter;SV:See dlingvigor;RSI:Rankingofsaltinjuryatseedlingstage;SFW/DMW:Theratioofshootfreshweightanddrymatterweightattheseedling;SW:Straw weightatmaturestage;SSR:Seedsettingrate;MPL:Mainpaniclelength;PH:Plantheight;NSMP:Numberofspikeletsinmainpanicle;ASDS:Averag esurvivaldaysofseedling;ISSVF:Theindividualofthesum ofscoresforvigorandfertility5.2信号分子传导植物的耐盐碱性是一个胁迫诱导和分子信号传导过程,该过程包括植株对外界胁迫的响应,胁迫分子信号的产生,识别与传导等几个环节,而植物耐盐}))"",........,.334期祁栋灵等:水稻耐盐碱性生理和遗传研究进展49l 碱相关基因是受盐碱胁迫诱导产生的信号调节. Hashimota等在盐胁迫下对2周龄稻苗的根系研究表明,水稻根中一种特异性诱导蛋白RsOsPR10的诱导表达不是通过脱落酸或水杨酸信号传导途径,而是通过茉莉酸信号转导途径.盐胁迫产生的信号能够诱导抗氧化酶基因的表达,进而启动抗氧化的微型调节系统,维持细胞环境的氧化还原动态平衡,从而保护细胞内各个器官组织免遭离子破坏.真核生物转录启始因子5A(IFSA)是一种启动胁迫信号转导中的级联放大反应,最终使细胞产生对胁迫的抗逆响应的胞质蛋白.在水稻中已发现两种转录启始因子,即OseIFSA一1和OseIF5A一2,受盐胁迫的诱导调控.一些学者对类黄酮的生物合成与水稻非生物逆境胁迫调节的分子基础之间的关系研究表明,编码水稻盐胁迫蛋白结构基因OSD一和OSAns的诱导表达受转录因子OSC1一MYB介导,同时,在OSDfr和OSAas基因的5端上游区发现了数个调控区,并认为类黄酮信号传导途径可能是逆境胁迫分子信号传导的重要途径之一.6水稻耐盐碱性基因克隆及转化近年来,借助分子生物学方法和技术,在水稻耐盐碱基因源的鉴定,重要基因的分离,克隆和转导等方面取得了较大进展.利用cDNA微阵列和RNA印迹技术在水稻中检测到57个高盐胁迫下诱导表达的基因,并定位和克隆了一些与耐盐性状有关的基因".Anoop等成功地把P5CS(2一氢吡咯5一羧酸合成酶)基因转人水稻中,并发现转基因水稻植株表现为较好的根生长和较高的生物量.Katsuhara等把大麦质膜上的一种水通道蛋白编码基因转入水稻并使其在转基因水稻中过量表达, 结果使水稻的茎根比率显着提高,对盐胁迫的敏感性下降.李南羿等用基因枪法将耐盐基因OPBP1转人水稻中,结果表明,在同等盐胁迫条件下转基因株系生长较快,叶绿素含量和生物学产量都显着高于未转基因对照.密穗野生稻(P.coarctata,2n=48)能忍受较长时间的海水浸泡,具有较强的耐盐性(30~40ds/m,Ece),利用远缘杂交,胚培养和生物技术等方法,将该野生稻的耐盐基因转人栽培稻中,获得了耐盐性较强的杂交后代.为了进一步了解转基因耐盐水稻后代植株的耐盐性表现,一些学者在获得转基因耐盐水稻植株后,又对其后代开展耐盐性研究,结果表明,第3代的转P5CS(吡咯啉一5一羧酸合成酶)基因水稻在高盐浓度胁迫下,根茎生长具有极高的耐盐性.尽管在水稻耐盐基因发掘和转化方面取得以上诸多进展,并且一些转基因耐盐碱水稻品系已进人环境试验或生物安全试验阶段,但由于还有一些关键性的问题尚未最终解决,还没有真正的转基因耐盐碱水稻品种应用于大田生产.7展望与建议7.1积极开展水稻耐盐碱种质资源的发掘与利用水稻的耐盐碱能力主要取决于种质自身的遗传特性.目前,耐盐碱水稻育种没有较大突破的原因之一就是当前所利用的耐盐碱亲本的遗传背景比较狭窄,类型较单一.将室内和室外鉴定试验相结合, 从现有保存资源中积极挖掘具有强耐盐碱能力的水。
【国家自然科学基金】_叶色突变体_基金支持热词逐年推荐_【万方软件创新助手】_20140802
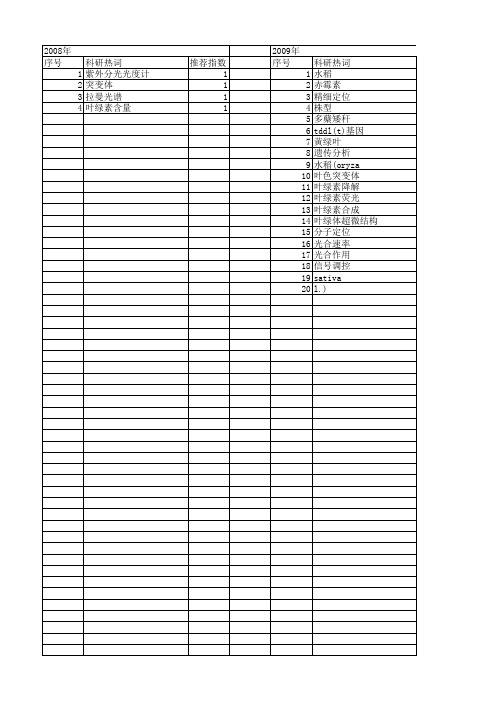
科研热词 推荐指数 水稻 5 白化转绿 2 白化 2 多分蘖矮秆 2 克隆 2 hw-1(t) 2 黄叶 1 遗传分析 1 精细定位 1 窄叶 1 温敏感 1 水稻(oryza sativa l.) 1 影响因素 1 定位 1 基因定位 1 基因克隆 1 叶色突变基因 1 叶色突变体 1 叶绿素 1 分子定位 1 光合作用参数 1
推荐指数 3 2 2 2 2 2 1 1 1 1 1 1 1 1 1 1 1 1 1 1
2010年 序号 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15
科研热词 水稻 叶色突变体 糖 白化突变体 玉米(zeamaysl.) 温敏感 拟南芥 基因定位 叶绿素代谢 叶绿素 mutator转座子 mu-tail-pcr lls突变体 indel标记 abc转运蛋白
2013年 序号 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25
科研热词 水稻 基因定位 叶色突变体 遗传分析 温敏感 黄绿叶突变体 黄叶突变体 超微结构 表达分析 联乙烯还原酶 突变体hfa-1 白化转绿 灰白叶 水稻(oryzasatival.) 失绿突变体 基因表达分析 图位克隆 叶绿素缺失 叶绿素含量 叶绿素 叶绿体 分子标记 光合色素 低温白化转绿 dvr
推荐指数 7 6 3 2 2 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1
2014年 序号 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16
2014年 科研热词 水稻 叶色突变体 黄绿叶突变体 遗传分析 紫花苜蓿 突变体库 基因 呈色机制 叶绿素荧光 叶绿体超微结构 变异性状 半致死剂量 候选基因分析 中华金叶榆子代苗 oscao ems(甲基磺酸乙酯) 推荐指数 2 2 1 1 1 1 1 1 1 1 1 1 1 1 1 1
中国水稻功能基因组研究进展与展望_肖景华

摘要
功能基因组研究是植物生命科学研究的核心领域之一. 从少数基因的克隆到重要农
关键词
功能基因组 全基因组 SNP 芯片 4D 基因组 水稻育种
艺性状的功能基因组解析, 我国水稻功能基因组研究实现了跨越式发展, 阐明了水稻育种 中的一些重大生物学问题 , 功能基因组的研究为水稻品种改良和育种技术变革奠定了基 础. 着眼未来, 我国科学家提出了继续推进“水稻2020”研究计划, 适时启动水稻4D基因组 的发展建议.
华中农业大学生命科学技术学院, 作物遗传改良国家重点实验室, 武汉 430070 * 联系人, E-mail: xiaojh@ 2015-04-14 收稿, 2015-05-04 接受, 2015-05-22 网络版发表 国家高技术研究发展计划(2012AA10A300)资助
引用格式 : 肖景华 , 吴昌银 , 袁猛 , 等 . 中国水稻功能基因组研究进展与展望 . 科学通报 , 2015, 60: 1711–1722 Xiao J H, Wu C Y, Yuan M, et al. The progress and perspective of rice functional genomics research in China (in Chinese). Chin Sci Bull, 2015, 60: 1711–1722, doi: 10.1360/N972015-00391
[2]
型突变体库、 核心种质资源和高密度的基因表达谱芯 片等功能基因组研究平台 , 发掘和克隆了一批控制 重要农艺性状的具有自主知识产权的功能基因 , 在 重要农艺性状形成的分子网络解析方面取得突破性 进展 , 为水稻基因组选择育种奠定了坚实基础 .1Biblioteka 水稻功能基因组研究的技术平台
庄福星在水稻“健身栽培”上推广与应用

庄福星在水稻“健身栽培”上的推广与应用庄福星是安徽省农业科学院引进美国先进的有机矿化技术生产的高科技产品,它含有作物生长所需要的多种营养元素,特别是含有甲壳素、黄腐酸和兼备改良土壤、促进农作物生长双重效果的进口原料kom,在农作物应用上,能够平衡养分供给,增强作物抗逆(旱、涝、冻害等)能力,在水稻生产应用上,具有促进水稻壮苗和调节苗情转化的功能。
在水稻的各个生育期与杀虫剂、杀菌剂混合施用,能够提高水稻的综合抗性和抗逆能力,以达到水稻“健身栽培”的功效。
1庄福星的作用与机理庄福星的作用与机理表现:一是含有作物生长发育所必需的各种元素和抗倒伏因子——壳聚糖,以及对农作物生长有益的多种微量元素;其中有机和无机养分的矿化作用,不仅提供农作物生长所必需的原料,更主要的是在作物生长发育的关键时期,能让作物更好地协调吸收、综合利用作物所必需的各种营养元素。
二是含有黄腐酸。
关于黄腐酸在农作物上的作用,近年成为作物栽培学、植物营养学、植物病理学等学科研究的热点。
黄腐酸可以明显地提高农作物的抗旱能力和防冻能力。
其机理是有效地控制气孔蒸腾,减少水分蒸发,保持体内水分。
黄腐酸对防治作物病害,特别是对水稻烂秧、棉花枯萎病、黄萎病、西瓜枯萎病及各种作物病毒病有很好防治效果。
即使在风调雨顺的年份,使用黄腐酸也能使农作物增产。
2庄福星在水稻“健身栽培”上的应用技术首先要降低秧田播种量,培育多蘖壮秧;其次是科学运用好庄福星在水稻高产栽培上的应用技术。
应用庄福星具体技术:一是水稻播种前用庄福星进行拌种(30 ml原液拌5 kg已浸泡催芽的种子)以及苗床喷施(用庄福星100 ml对水200~300倍喷于苗床露籽和土壤上)。
二是喷施送嫁肥。
在水稻移栽前3~5 d用庄福星900 ml/hm2对水300~500倍作为“送嫁肥”叶面喷施。
三是在水稻拔节至齐穗期间,用庄福星900 ml/hm2对水300~500倍叶面喷施。
四是在施好倒2叶露尖期的保花肥和抽穗期前后的粒肥的基础上,同时叶面喷施庄福星。
草地早熟禾NADH-GOGAT基因的克隆及表达分析

草地早熟禾NADH-GOGAT基因的克隆及表达分析陈阳;孙华山;王玉书;张学通;师冉;熊良兵;金一锋【摘要】氮素是制约草坪草生长发育的重要因素之一,GS/GOGAT循环是植物氮同化的主要途径,谷氨酸合酶(glu-tamine:2-oxoglutarate amidotransferase,glutamate synthase,GOGAT)在植物氮素转化过程中发挥着重要的催化作用.本研究以草地早熟禾(Poa pratensis)为试材,基于二代测序RNA-seq相关数据,克隆得到草地早熟禾NADH-GOGAT基因,并进行生物信息学分析.结果表明:草地早熟禾NADH-GOGAT基因序列为3236 bp,完整的开放阅读框(Open Reading Frame Finder,ORF)为1338 bp,编码445个氨基酸残基组成的蛋白;预测NADH-GOGAT蛋白的分子量为49.89KD,等电点(isoelectric point,pI)为6.11,无信号肽,含有1个Glu-syn-central结构域,属于Glu-syn-central超级家族;二级结构以无规则卷曲、α-螺旋、延伸链和β-转角为主;同源进化分析结果表明草地早熟禾NADH-GOGAT与硬粒黑小麦氨基酸序列相似度为97%.草地早熟禾NADH-GOGAT基因在叶部的相对表达量高于根与茎,低氮浓度更有利于该基因的表达.NADH-GOGAT在干旱胁迫和水氮互作中表达差异显著(P<0.05).草地早熟禾NA D H-GOGA T基因的克隆,及相关序列分析和基因表达,对进一步研究该基因的功能和草地早熟禾GS/GOGA T循环的调控过程具有一定意义.【期刊名称】《草地学报》【年(卷),期】2019(027)002【总页数】7页(P459-465)【关键词】草地早熟禾;谷氨酸合酶;基因克隆;氮素调控【作者】陈阳;孙华山;王玉书;张学通;师冉;熊良兵;金一锋【作者单位】齐齐哈尔大学生命科学与农林学院,抗性基因工程与寒地生物多样性保护黑龙江省重点实验室,黑龙江齐齐哈尔161006;东北农业大学园艺园林学院,黑龙江哈尔滨150030;齐齐哈尔大学生命科学与农林学院,抗性基因工程与寒地生物多样性保护黑龙江省重点实验室,黑龙江齐齐哈尔161006;齐齐哈尔大学生命科学与农林学院,抗性基因工程与寒地生物多样性保护黑龙江省重点实验室,黑龙江齐齐哈尔161006;齐齐哈尔大学生命科学与农林学院,抗性基因工程与寒地生物多样性保护黑龙江省重点实验室,黑龙江齐齐哈尔161006;齐齐哈尔大学生命科学与农林学院,抗性基因工程与寒地生物多样性保护黑龙江省重点实验室,黑龙江齐齐哈尔161006;齐齐哈尔大学生命科学与农林学院,抗性基因工程与寒地生物多样性保护黑龙江省重点实验室,黑龙江齐齐哈尔161006【正文语种】中文【中图分类】S543草地早熟禾(Poa pratensis)作为优质的冷季型草坪草,凭借良好的坪用性状及耐寒性广泛应用于我国北方的园林绿化[1]。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
生命科学Chinese Bulletin of Life Sciences第19卷 第6期
2007年12月Vol. 19, No. 6
Dec., 2007
文章编号 :1004-0374(2007)06-0614-02
进一步提高水稻产量,最大限度满足国家对食物安全的需求是水稻遗传育种的重大任务。水稻干物质产量的90%-95%来自光合作用。目前高产水稻光能利用率也仅1.5%-2.0%。理论上,植物光能利用率可达13.0%-14.0%,水稻理想的光能利用率应达3.0%-5.0%[1]。因此,培育高光效的超高产品种是提高产量的主要途径之一。长期以来,水稻光合作用的相关研究大多停留在生理水平上,同时,传统的杂交育种手段在改良水稻光能利用方面至今尚未取得令人满意的结果,而叶色突变体是开展光合作用研究的理想材料。水稻黄绿叶突变体ygl1是水稻栽培品种镇恢249的自然突变体,该突变体前期叶片为黄色,中期慢慢转绿,后期叶色接近野生型。与野生型相比,ygl1突变体有较高光合效率和较强的耐受光抑制能力,因而仍能获得较高的产量(亩产900.12斤)。高等植物叶绿素的生物合成与叶绿体的发育对其能否进行正常的光合作用起关键作用[2]。叶绿体的许多蛋白复合体的蛋白和叶绿素生物合成所需酶类都是由核基因及质体基因编码。同时,叶绿素的合成与叶绿体的发育涉及核基因和叶绿体基因协同调控的复杂过程,其中一些基因的突变将造成叶绿素缺乏或叶绿体发育缺陷,导致光合作用受阻[3-5]。从黄绿叶突变体ygl1克隆与叶绿素合成与降解、叶绿体分化与发育、光合作用代谢途径相关的新基因,对开展光合系统结构、光合机构的应答及其调控机制研究有重要意义,也为水稻高光效生理研究和水稻黄绿叶突变体ygl1在生产上的应用提供了理论依据。在国家“863”、“973”计划以及国家自然科学基金等项目的支持下,南京农业大学水稻研究所、中国农业科学院作物科学研究所等单位的科研人员利用黄绿叶突变体ygl1为材料,通过数年研究,克隆出水稻黄绿叶突变基因ygl1,且从生理生化、细胞形态学等方面对ygl1突变体进行了比较系统的研究[6]。研究表明,YGL1基因编码叶绿素合成酶,催化叶绿素合成过程中最后一步叶绿素a的形成。核苷酸序列分析发现突变体与野生型在叶绿素合成酶基因的编码区上有单个碱基差异,由胸腺嘧啶(T)突变为胞嘧啶(C),造成所编码的脯氨酸(Proline)到丝氨酸(Serline)的改变,其具体研究过程如下:为了研究ygl1突变体的遗传特性,用ygl1突变体分别与PA64、W002、USSR5和02428等几个栽培品种构建了四个正反交杂交组合,结果所有的F1
显示野生型绿苗表型,后代F2出现黄、绿苗分离,
分离比接近3∶1,经χ2测验(χ2<χ2
0.05=3.84;P>
0.05),符合3∶1分离,表明ygl1的黄绿叶表型由一对隐性核基因控制。利用已公布图谱上SSR分子标记和ygl1突变体与培矮64杂交组合衍生的F2代临时性分离群体的252个突变体单株对突变基因进行初步定位[7],把突变基因定位于第5染色体上SSR标记RM516和RM164之间,遗传距离分别为4.8 cM和13.1 cM;随后通过扩大定位群体,并根据水稻基因组数据开发新型分子标记[8],构建了ygl1基因区域高密度遗传图谱和细菌人工染色体重叠群,最终将突变基因精细定位于细菌人工染色体AC136221上大约11kb区间内,与基于PCR的剪切扩增多态性(cleavedamplified polymorphicsequences,CAPS)标记P25、P26共分离。利用基因分析与预测软件对定位的11 kb区域进行预测,发现仅有两个开放阅读框(openreading frames, ORFs) :一个为叶绿素合成酶基因(chlorophyll synthetasegene)[9-13] ,我们把它称为
水稻黄绿叶基因的克隆及应用吴自明1,张 欣2,万建民2
*
(1南京农业大学作物遗传与种质创新国家重点实验室,江苏省植物基因工程中心,南京 210095;2 中国农业科学院作物科学研究所,北京 100081)
收稿日期:2007-07-10;修回日期:2007-07-17基金项目:“973”国家重点基础研究发展计划(2006CB1017000;2006CB100201) ;“863”国家科技攻关计划(2006AA100101) ; 国家自然科学资金资助项目(30571142) ; 长江学者和创新团队发展计划作者简介:吴自明(1974—),男,博士,讲师;万建民(1960—),男,博士,博士生导师,*通讯作者,E-mail:wanjm@caas.net.cn
·研究快讯·615第6期吴自明,等:水稻黄绿叶基因的克隆及应用
YGL1基因;另一个为em基因(embryogenicabscisiacid-inducible gene)。通过RT-PCR分别扩增野生型和突变体两个开放阅读框编码区cDNA,测序后分析发现野生型和突变体的em基因cDNA没有任何差异,仅在叶绿素合成酶基因YGL1的编码区cDNA上有单碱基差异,由胸腺嘧啶突变为胞嘧啶,造成脯氨酸到丝氨酸的改变。因此,我们把YGL1基因确定为ygl1的候选基因。进一步构建了含有YGL1编码区cDNA双元表达载体,通过农杆菌介导的方法将此基因转化到具有ygl1等位基因黄绿叶表型的粳稻品系黄叶粳,恢复绿色表型的植株经PCR检测、Southern blot分析、T1代分离实验及恢复表型植株叶绿素含量分析,证明YGL1就是ygl1突变体的突变基因。YGL1以单拷贝的形式存在于水稻基因组中,编码叶绿素合成酶,催化叶绿素酸酯植醇化,生成叶绿素a,这一合成步骤对叶绿素a辅基蛋白的翻译和积累,类囊体膜组分的稳定装配起重要作用[14]。氨基酸序列同源性和进化树分析表明,不同物种叶绿素合成酶基因有很强的保守性,水稻叶绿素合成酶YGL1与同为禾本科植物燕麦的叶绿素合成酶比其他生物有更近的亲缘关系。体外原核表达重组蛋白酯化活性分析显示,突变基因重组蛋白ygl1较野生型YGL1重组蛋白酯化活性下降,表明脯氨酸到丝氨酸的改变,导致叶绿素合成酶活性下降;ygl1突变体的叶绿素积累速率减慢和叶绿素合成中间产物的积累也充分表明叶绿素合成酶催化活性下降;YGL1具有组成型表达特性,单碱基突变并没有影响到突变体体内的表达水平,然而,与叶绿素合成和叶绿体发育相关的基因表达受到了不同程度的影响,尤其是编码叶绿素a/b结合蛋白cab1R基因在苗期叶片中的表达受到强烈抑制,这些结果为叶绿素或叶绿素中间代谢产物水平反馈调控编码叶绿体相关核基因表达提供了直接的证据。叶绿素在光合作用的光能吸收、传递和转换中起着关键作用。早期研究认为,在一定范围内,叶绿素含量与光合速率成正相关关系。水稻在经济器官形成和充实期,剑叶叶绿素含量保持在较高水平,能使叶片保持较高的光合速率。水稻黄绿叶突变体ygl1,其叶绿素b含量适量减少, 降低捕光色素的比例,但是提高光合作用的活性, 同时降低在高光强下的光抑制现象。该突变体抽穗前叶绿素含量与野生型没有显著差异,这是突变体仍能获得较高产量的主要原因。水稻黄绿叶突变体ygl1叶绿素含量适当减少,光合结构对强光的耐受性升高,最大光合速率显著上升,所以水稻黄绿叶突变体ygl1不仅可用于叶绿素生物合成、叶绿体分化与发育和水稻高光效生理的研究等基础研究,其黄绿叶特性还可作为标记性状在杂种优势利用中应用,如在良种繁育和杂交育种实践中,可以通过回交转育或转基因的方法,将ygl1黄绿叶基因导入光(温)敏两用核不育系当中,利用叶色变异作为标记性状,不但可以测定水稻种子纯度,大大降低田间检测种子纯度的成本和时间,还可在苗期去杂,克服光(温)敏核不育系在制种时常见的由于温度变化而造成育性恢复,导致杂种F1纯度不同程度的下降所带来的杂交稻产量下降的损失,对简化杂交稻纯度鉴定手续、节约纯度鉴定时间和费用及提供杂交稻生产的田间纯度具有重要意义。[参 考 文 献][1]Loomis R S, Williams W A. Maximum crop productivity: anestimate. Crop Sci, 1963, 3: 67-72[2]Eckhardt U, Grimm B, Hortensteiner S. Recent advances inchlorophyll biosynthesis and breakdown in higher plants.Plant Mol Biol, 2004, 56: 1-14[3]Falbel T G, Staehelin L A. Partial block in the early steps ofthe chlorophyll synthesis pathway: A common feature ofchlorophyll b-deficient mutants. Plant Physiol, 1996, 97:311-320[4]Jung K H, Hur J, Ryu C H, et al. Characterization of a ricechlorophyll-deficient mutant using the T-DNA gene-trapsystem. Plant Cell Physiol, 2003, 44: 463-472[5]Tanya C F, Staehelin L A. Characterization of a family ofchlorophyll-deficient wheat (Triticum) and barley (Hordeumvulgare) mutants with defects in the magnesium-insertionstep of chlorophyll biosynthesis. Plant Physiol, 1994, 104:639-648[6]Wu Z M, Zhang X, He B, et al. A chlorophyll-deficient ricemutant with impaired chlorophyllide esterification inchlorophyll biosynthesis, Plant Physiol, 2007, 145: 29-40[7]McCouch S R, Teytelman L, Xu Y B. Development andmapping of 2240 new SSR markers for rice (Oryza sativaL.). DNA Res, 2002, 9: 199-207[8]Thiel T, Kota R, Grosse I, et al. SNP2CAPS: a SNP andINDEL analysis tool for CAPS marker development. NucleicAcids Res, 2004, 32: e5[9]Lindsten A, Welch C J, Schoch S, et al. Chlorophyll syn-thetase is latent in well-preserved prolamellar bodies of eti-olated wheat. Physiolgia Plantarum, 1990, 80: 277[10]Lopez J C, Ryan S, Blankenship R E. Sequence of the bchGgene from Chloroflexus aurantiacus: relationship betweenchlorophyll synthase and other polyprenyl transferases. JBacteriol, 1996, 178: 3363-3373[11]Oster U, Bauer C E, Rüdiger W. Characterization of chloro-phyll a and bacteriochlorophyll a synthases by heterolo-gous expression in Escherichia coli. J Biol Chem, 1997, 272:9671-9676[12]Oster U, Rüdiger W. The G4 gene of Arabidopsis thalianaencodes a chlorophyll synthase of etiolated plants. Bot Acta,1997, 110: 420-423[13]Schmid H C, Oster U, Kögel J. Cloning and characterisationof chlorophyll synthase from Avena sativa. Biol Chem,2001, 382: 903 -911[14]Schmid HC, Rassadina V, Oster U. Pre-loading of chloro-phyll synthase with tetraprenyl diphosphate is an obliga-tory step in chlorophyll biosynthesis. Biol Chem, 2002,383: 1769 -1776