32-第13章 基因组进化的模式-基因水平的进化
基因组的进化

OH
5´
3´ OH
RNA一级结构
当前您正浏览第十二页,共二十四页。
OH
第二节 基因组进化的模式
1.遗传系统的起源
1.2基因组的起源
RNA世界向DNA世界的过渡可以概括为:地球上最早出现的生物 大分子为RNA,RNA同时具有催化与编码两种功能。RNA可以催化 肽键形成并合成蛋白质,此后RNA与蛋白质联手以RNA为模板合成 DNA。这是一个关键的转变时期,生命世界的三大主要多聚分子, RNA、蛋白质和DNA的分工基本定形:RNA的编码功能由DNA取代, 催化功能转移到蛋白质,RNA自身则称为传达遗传信息的中介分子。
初始RNA分子
新互补拷贝
早期RNA世界RNA分子的拷贝
当前您正浏览第十页,共二十四页。来自第二节 基因组进化的模式
1.遗传系统的起源
1.2基因组的起源
核苷酸 TA C T
GTAC TCG
还原
脱氧核糖核苷酸 dA dC dG
dT dC dA dC dG dA
RNA拷贝
第一个拷贝的DNA分子
RNA
DNA
编码RNA分子转变为第一个DNA分子
成新链。由此产生的子代DNA中,会将这一变异一直保留下去。
当前您正浏览第二页,共二十四页。
第一节 基因组进化的分子基础
1.突变
1.1突变的效应 ➢突变对基因组的影响
同义突变 错义突变 终止突变 连读突变 移码突变
➢突变对多细胞生物的影响
功能丧失突变(loss-of-function mutation):通常是一个减弱或消除蛋白质活性的突 变造成的结果。多数功能丧失突变是隐性性状。
当前您正浏览第二十四页,共二十四页。
2.3双链断裂重组模型
进化中的基因组演化

进化中的基因组演化基因组演化是生物进化的重要方面,它涉及到基因组的复制、突变、选择以及遗传流动等过程。
通过这些演化过程,生物的基因组在时间的推移中发生着变化和适应,以适应环境的改变和生存的需求。
本文将探讨基因组演化的基本原理和其在进化中的作用。
1. 基因组的复制和突变基因组的复制是基本的遗传过程之一,它使得一个个体可以将其基因组传递给下一代。
然而,基因组复制并非完全准确,会产生突变。
突变是基因组演化的核心机制,它导致基因组中的遗传信息发生改变。
突变可以是点突变、插入或删除等不同形式,从而导致基因型和表型的变化。
2. 自然选择和基因组的适应性自然选择是基因组演化中的另一个重要驱动因素。
它通过筛选与环境适应性更强的个体,使得其基因组在群体中逐渐变得更加普遍。
这种适应性的积累使得物种能够更好地适应其生存环境,并提高其生存和繁殖的机会。
3. 遗传流动和基因组的多样性遗传流动是指不同个体之间基因信息的交流和共享。
它可以通过基因的迁移、杂交和基因池的变化等方式发生。
遗传流动可以增加基因组的多样性,从而使得物种更具适应性和生存力。
然而,过度的遗传流动也可能导致基因组的同质化和物种的退化。
4. 基因组重组和变异积累基因组重组是指在有性生殖中的染色体交换和基因片段重组。
它使得不同的基因片段能够重新组合,从而产生新的基因组组合和变异。
这种基因组重组和变异的积累是基本的演化过程之一,它增加了基因组的多样性和变化速度。
总结起来,基因组演化在生物进化过程中起着重要的作用。
通过基因组的复制、突变、自然选择、遗传流动、基因组重组和变异积累等过程,生物能够适应环境的变化和生存的需求。
基因组演化是生物多样性的来源之一,同时也是人类对进化过程的深入研究的重要内容之一。
通过对基因组演化的研究,人们可以更好地理解物种的起源、形态的多样性以及生物的适应性进化。
此外,对基因组演化的理解还可以为医学和农业领域的研究提供重要的参考,例如通过对基因组的分析和比较,可以揭示人类疾病的遗传基础,以及改良农作物的育种方法。
基因组与进化

15
2019/12/27
CB
(1)碱基切除修复
由糖苷键酶启动,可 修复受损碱基,AP位 点,和DNA断裂
16
2019/12/27
CB
(2)核苷酸切除修复
E.coli的切除修复机 制:UvrABC酶复 合体),12bp
17
2019/12/27
CB
(3)错配切除修复
识别位点:DNA 复制与子链甲基 化的时间差; E.coli有长、短、 极短链修复系统, 由不同的酶复合 体负责
5)基因冗余并非真正的多余
54
2019/12/27
CB
三、非编码序列的扩张
非编码序列的作用 1)非编码序列可能具有某种未知的功能: 染色质基质附着区(植物MITE)、转录后调控(3’-UTR,MIR)、基 因表达的多样性(转座子) 2)非编码序列可能是”自私DNA” 1、转座子和基因组进化 1)最重要的作用是引起基因重排,有利有弊。 2)其插入可改变临近基因的表达模式或剪接模式(图)
很难找到相关证据,主要集中在逆转录病毒和转座成分.
53
2019/12/27
CB
4、基因冗余
1)基因冗余与进化
2)类型(进化分类)
(1)年轻的重复基因(2)正在向新功能基因过渡的重复基因 (3)保留部分功能重叠的重复基因
3)看家基因很少重复,与发育有关的或涉及生物多样性的, 特别是多功能域基因常见
4)自然选择有差别地淘汰重复基因,与重复基因的功能及 对生物进化所具有的潜在意义有关
CB
52
2019/12/27
CB
3、DNA水平转移
1)原核基因的DNA水平转移
可能在原核生物的分化与种属的形成过程中发挥了作用.
32-第13章 基因组进化的模式-基因水平的进化

HOX基因簇的进化
Hox基因家 族在动物的 发育与体节 模式确定中 起着极其重 要的作用。 哺乳动物的 4个Hox基因 簇的来源可 以追溯到原 始的HoxEvx/Meox基 因的加倍, 趋异与缺失, 代表基因进 化的一种主 要方式。
基因重复与适应性进化球蛋白基因
哺乳动物现存球蛋白基因 经历过前后5次加倍与趋 异。原和原球蛋白的 祖先基因在约5亿年前加 倍,在2亿年之后球蛋 白和球蛋白祖先基因又 出现加倍,随之各自发生 分离。现存的肌球蛋白 和球蛋白的功能发 生了 趋异,适应了生物进 化的 需要: 胎儿期表达的球蛋白可满 足从母体血液吸收氧的功 能,成年表达的球蛋白与 氧结合能力减弱。
有弊也有利
人类 颌肌 肌球 蛋白 基因 的 突变
基因组点突变包括碱基代换与增减(indel, insertion/ deletion ), 基因编 码区和基因调控区如发生点突变,会影响基因编码功能与基因的表达调 控。这些突变有些是有害的,有些是有利的。如血红蛋白基因翻译相关 kozak序列的碱基代换产生地中海贫血病,颌肌肌球蛋白基因碱基缺失 造成移码突变促使人类大脑体积的扩增。
β-球蛋白基因
加倍趋异
哺乳动物和禽类β-球蛋白 基因有着共同的祖先。 原 始β-球蛋白基因( pβ1和 pβ2 )和调控顺序(LCR和 增强子)在经历一次加倍 后,分为两支进化:一支 向真哺乳亚纲和袋鼠发展; 一支成为禽类现存β-球 蛋白基因(p, Ε,pβA和 pβB )的先祖。 PNAS 98:1327-1329, 2001
蛋 白 质 体 外 进 化
体外蛋白质进化的设计原理类似于DNA洗牌。利用嵌合 寡聚DNA引物扩放靶基因或同源基因的外显子,随 后将扩放产物混合,变性后复性,再完成自身引物 PCR。最后通过实验筛选重组基因,检测蛋白活性。
31-第13章 基因组进化的模式-基因组水平的进化

海胆重复顺序。
DNA水平转移--
原核生物进化的主要方式
原核生物的进化途径主要是DNA的水平转移,即通过摄取 外源基因改变基因组的组成以适应自然选择。
在超嗜热菌Hyperthermophilic bacterium) ,产液菌 (Aquifex aeolicus)和詹氏甲烷球菌之间,螺旋菌 (Helicobacter pylori)和不同远缘细菌之间均有基 因交流现象。
PNAS 104: 8397–8402, 2007
大多数植物基因组均为多倍体
1) 大多数植物基因组均由同源多倍体和异源 多 倍体组成, 是基因数目增加的主要方式。
2) 玉米染色体组由染色体基数n=5加倍为n=10 。 小麦为6倍体(含A, B, D=7, n=21), 为异源多 倍 体。油菜为异源多倍体(n=19), 棉花, 烟草 为 多倍体, 香蕉为3倍体。
能量促使菌毛收缩,使附
着的外源DNA的一条单链 通过受体蛋白EA进入膜通 道EC,同时另一条单链被 核酸酶降解。解旋酶和 DNA转移酶(FA)与单链 DNA结合蛋白(ssb)和RecA
联手使进入细胞内的外源
单链DNA与受体DNA接触 ,然后RecA蛋白质与单链 DNA组成的蛋白核酸纤丝 与受体DNA发生同源重组
脊椎动物基因组2R假说
2R假说认为,在无颌类脊索动物(jawless vertebrate)出现之前和出现之后分别出现过 一次全基因组的加倍,即脊索动物有过两轮全 基因组加倍。
Genome Research 11:667–670 , 2001 2008年6月完成文昌鱼的基因组序列测序。对文
昌鱼和脊椎动物基因组中保留下来的17个先祖 脊索动物连锁群进行染色体虚拟重建。 结果证 实在有颌类脊索动物演化过程中, 确实发生了 两轮完整基因组加倍现象,2R假说是正确的。 Nature 453, 1064-1071, 2008
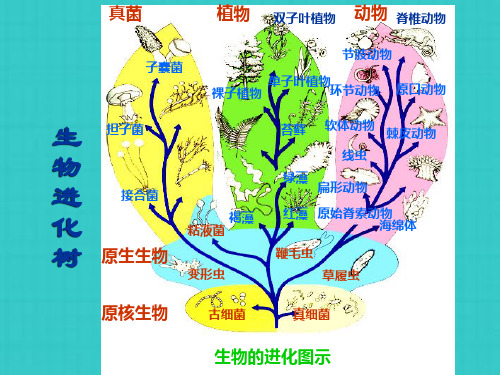
基因组进化的模式
斯卑尔脱小麦 T. spelta(AABBDD)
面包小麦 T. aetivum(AABBDD)
单个基因以及染色体区段加倍在进化过程中经常出现
• 单个基因以及基因群加倍 在进化过 程中经常出现
– 多基因家族,同一家族的不同成员最后 均可追溯到最初的古老基因组的某一祖 先基因
• 加倍机制 不等交换
– 同源染色体不等交换:位于同源染色体不同 位置的相似核苷酸序列之间发生的重组事件
• 全套基因组的加倍 使基因数目急剧增加
– 同源多倍体:减数分裂发生差错时使配子体含 2套染色体(双倍体配子),融合产生同源多 倍体
不直接导致基因扩张,提供了产生新基因的可能
• 重复基因持续丢失 • 突变不会对存活力造成严重影响 • 突变往往会造成基因的失活,成为假基因 • 偶而有一些突变会使加倍的基因具有新的功能,
内含子如何能插入到完整而连续编码的基因序列中而对其功能丝毫无损?
细胞器,如喜氧细菌成为线粒体,蓝藻成为叶绿体。
重复区域跨越了50%的基因组 基因组加倍和种间基因转移的结合
用电击提供能量,从简单小分子可得到复杂大分子。
University of Colorado
Boulder, CO, USA
?
•核酸起源说
选择性的
2.2 基因重排
• 具创新功能的蛋白质更多是通过现有基因的 重排产生,功能域区段重排。
外显子的特点:
1、外显子形成的多肽链独立结构域
2、内含子的位置通常在两个相连的独立结构或功 能的多肽链编码序列之间
自发多聚体化
初始RNA分子
新互补拷贝
早期RNA世界RNA分子的拷贝
地聚合。这一过程很不精确,但可产生许多RNA序列。
水平进化名词解释
水平进化名词解释
水平进化是指生物在种群内演化出不同的性状,而这些性状并没有改变生物在分类学上的地位。
以下是五个水平进化的名词解释及例句:
1. 基因漂变:指在一个种群中,某些基因的频率由于随机抽样误差而发生的变化。
例如,在一个种群中,某个基因的频率可能会因为偶然事件而增加或减少。
例句:基因漂变导致了某些种群中特定基因的消失。
2. 基因流动:指基因在种群之间的迁移。
基因流动可以通过个体的迁徙、交配等方式实现。
例如,当一个种群中的个体与其他种群的个体交配时,基因就会在种群之间流动。
例句:基因流动有助于维持种群的遗传多样性。
3. 中性进化:指在自然选择对某些基因的影响不明显时,这些基因的频率会随着时间的推移而发生随机变化。
中性进化不是由自然选择驱动的,而是由遗传漂变和基因流动等随机因素引起的。
例句:中性进化导致了一些不影响生物生存和繁殖的性状的变化。
4. 趋同进化:指不同的物种在相似的环境条件下,独立地演化出相似的适应性特征。
这些相似的特征并不是由于共同祖先遗传下来的,而是由于相似的自然选择压力导致的。
例句:鲨鱼和海豚在外观和行为上有很多相似之处,这是趋同进化的结果。
5. 辐射进化:指一个物种在短时间内快速演化出多个不同的物种,形成一个辐射状的分类群。
这种进化通常发生在物种进入新的环境或生态位时。
例句:灵长类动物在进化过程中经历了辐射进化,形成了多种不同的物种。
这些名词解释及例句展示了水平进化的不同方面,包括基因频率的变化、基因流动、中性进化、趋同进化和辐射进化。
生物科学中的基因组进化和基因功能调控
生物科学中的基因组进化和基因功能调控基因组的进化是生物学研究的重要方向,它研究基因组的演化规律以及不同物种的基因组之间的异同。
基因组由DNA分子组成,是生命体的遗传信息存储库,基因组的演化过程涉及到分子层面的变化、基因复制、基因组再组合和拼接以及自然选择等多个方面。
基因组的进化是通过基因改变和多样性的消失和积累来实现的。
基因组演化的过程中,基因不断地经过复制、突变和自然选择,不断形成新的差异和多样性。
生物进化的一个重要方面是基因组重构,包括分裂、重组、倒位和复制等形式。
进化过程中还涉及基因家族的出现和消失,基因转移等重要事件。
基因组进化的研究有助于我们理解基因序列演化的机制和不同物种之间的关系。
例如,研究化石遗传学可以通过古代DNA重建不同物种的基因组,为我们了解物种演化提供了更为直接和具体的证据。
另外,随着遗传学技术的进展,每个人的基因组也可以更为精确地测序和比较,这在疾病预防和治疗等方面也有很大的应用价值。
例如,遗传性疾病的治疗方法就可以根据个体基因组变异的情况来制定。
基因功能调控是指生物体内各种分子的相互作用,调节着基因的表达,以实现生命体适应环境和维持稳态。
基因表达调控主要由转录因子和非编码RNA来实现。
确定基因调控机制对于理解生物体的生理生化过程具有至关重要的意义。
转录因子是一类可以调节基因表达的蛋白质,对基因转录起着关键作用。
在基因表达调控的过程中,转录因子直接与DNA结合,可以在某些区域上引入修饰,如甲基化和去甲基化,从而影响基因的转录起始。
非编码RNA与转录因子一样也在基因表达调控中起着重要作用。
非编码RNA是指不编码蛋白质的RNA分子,但是它们可以通过抑制或促进基因表达来起到调控作用。
近年来,许多新型的非编码RNA已经得到发现,而发现这些RNA的功能注解需要进一步的实验验证。
基因功能调控是生物进化的一个重要方面。
基因调控的机制在不同物种间存在着很大的异同。
例如,在人类和其他物种的基因组中,同样拥有某一个基因,但是它们在实际功能上可能大不相同。
基因进化的机制与模式
基因进化的机制与模式基因是生命的基本单位,是控制个体遗传特征的基础。
基因进化是基因在物种中随着时间的推移自然选择、随机漂变、基因流动等自然力量作用下的演化过程。
基因进化有其自身的机制与模式,探究基因进化的机制与模式有利于我们了解生命中的基本规律。
I. 自然选择自然选择是指环境作为自然力量对生物群体中表现出较强的遗传特征的个体进行筛选的过程。
在维护环境稳定的情况下,自然选择可以促进生物体适应环境的进化。
例如,当一种植物生长在隆起的山地中,面对气候变化和山地地形的困扰,它很有可能具备更加强壮的根系、能够更好的抵抗自然环境的能力,这样它就可以在远离山陵地区,如平地,生长更为迅速和成功。
II. 随机漂变随机漂变指的是基因频率突然变化的随机现象。
随机漂变是用于说明一些相对孤立的群体中基因变化的力量。
例如,一些地形相对孤立、环境异常困难、数量比较少的植物和动物,也面临艰难的生存环境和基因漂变的现象。
III. 基因流动基因流动是指两个或多个不同的物种间,当它们互相交配并产生后代时,基因从一个物种流入另一个物种的过程。
基因流动可以通过遗传杂交、基因转移、基因重组等方式实现。
例如,当小羊妈妈和大羊爸爸交配时,基因从大羊中流入小羊中,这将改变小羊的遗传特征,随后小羊将表现出部分大羊的遗传特征,这对生物体的进化有重要意义。
IV. 分岔进化分岔进化是指一种物种在进化上的分裂,产生了两个或多个不同的进化路线或亚种群,从而在不同方面的遗传特征上出现了差异。
例如,一些动物在进化的过程中分成了两种亚种群,对于这两种亚种群的交配,随后将出现两种不同的遗传特征,对于二者的个体可有区别的深刻影响。
总之, 基因进化的机制与模式是一个复杂而全面的过程,涉及许多自然力量的影响,然而要彻底理解这些机制,必须从基因的角度来研究它们。
通过学习基因进化的机制与模式,我们可以更好地理解自然的生命规律,并在保护生态环境和人类健康的方面做出更为明智和有效的决策。
遗传学中的基因进化过程
遗传学中的基因进化过程从我们的爷爷奶奶那一代开始,我们的身体内就开始了基因遗传。
随着我们的父母的精子和卵子的结合,我们的基因组成开始慢慢建立起来。
然而,基因并不是静态的存在,它们会在进化和适应环境的过程中,经历复杂的变化。
在遗传学中,基因进化是指基因在进化中的改变和演化过程。
它不仅关系到个体的繁殖和适应性,也直接影响到物种的生存和繁衍。
基因进化在一定程度上也解释了为什么不同物种之间,拥有不同的外貌和行为特点。
基因进化的过程可以从以下几个方面进行解释和说明。
一、随机突变随机突变是基因进化的首要原因。
它指的是基因随机发生的突变、缺失、或者复制错误,也可以通过突变新增加一些基因序列。
虽然随机突变的概率非常低,但由于人口数量庞大,每个个体在自我复制遗传时的不完全复制引起的差异导致了漫长的遗传演化过程。
这就是为什么在基因进化的角度上,人的出现算得上是非常不容易的。
二、自然选择自然选择是自然界中治理基因进化的最关键因素之一。
生命物种总数目是一个相对稳定的数量。
而生命体之间存在的竞争和生存压力,导致了某些个体表现更好,更有能力适应生存环境而生存下来的生存方式。
这些个体遗传到的基因自然也就更多的流传给下一代,因此基因的进化也因此受到了影响。
三、基因漂变基因漂变是指一些随机的因素,例如地震、火山爆发、旅行等,引起基因在一个地理范围内随机变异的现象。
由于基因交流受到阻碍和约束,就会发生与人口数量相对较少的群体中的基因存在很大的随机变化,导致漂变。
这种现象特别多地出现在岛国或山区。
四、基因流动基因流动指的是一个个体或种群的基因大量从一个群体流入另一个群体的过程。
在基因流动方面,虽然事件较为罕见,但最终结局是有可能改变一个物种群体的遗传构成的。
这也是为什么遗传学会需要对人类进行更加深入的研究。
综上所述,基因进化是不可避免的,它一直在影响着我们身体内的遗传基因。
虽然这个过程在短时间内相对较慢、较难察觉,但长时间的积累和堆积,定会让我们发现基因进化的重要性和影响力。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
外显子洗牌
外显子洗牌(Exon shuffling )系指产生新基因的一 种分子机制。来自不同基因的两个或多个外 显子,经过异位(ectopiclly)重组或加倍形成 新的具有外显子-内含子结构的基因。
外显子洗牌的依据:1)外显子编码的多肽链往 往形成一个相对独立的结构域(module); 2)内含子处独立的结构域之间;3)氢键或 二硫键出现在外显子编码多肽链内部;4)重 复的外显子多肽链产物和重复的结构或功能 单元对应;5)非同源基因同源外显子编码相 似的多肽结构。
基因的进化速率是不相同的
1)生物信息学研究发现,与基础代谢有关的基因相对比 较保守,进化速率较慢;调控基因涉及表达模式的确 定,进化速率较快;
2)涉及环境适应性的基因进化速率相对较快。人类和黑 猩猩的进化树中,与免疫、细胞凋亡和精子发育相关 的基因演变的速度甚至比随机突变还要快。因为病毒 和其他病原体的进化速度很快,人类免疫系统不断受 到新出现的病毒的威胁,因此,与抵抗疾病相关的基 因进化更加迅速。
第13章-2 基因水平的进化
新基因的产生
新基因产生的主要方式: 1) 基因加倍之后的趋异,是新基因产生的主要方式。 2) 结构域洗牌, 即不同的结构域加倍或重组, 产生创新
功能基因。真核生物约19%的基因产生于外显子洗 牌。 3) 基因裂变与融合, 一分为二或者合二为一。原核生物 约0.5%的基因由此产生。 4) 嬗变, 由非编码顺序转变为编码顺序(例罕见子)。
定向 DNA 洗牌
不同生物的同源基因编码序列仅有少量的差异,但编码的蛋白产 物在功能上有很大差别。将这些同源基因的DNA混合,然后将 其断裂为长短不一的片段。解链后再复性,可形成杂合双链, 再完成自身引物PCR,形成经洗牌重组的DNA。从这些重组 DNA克隆中可筛选到蛋白活性改进的基因。通过这种方法,已 将头孢菌素酶(cephalosporinase)的活性提高250倍。 Nature, 1998, 391: 288-291
Hoxc-6
基因在 鸡和蛇 胚胎中 的表达 模式
Hox 基因决定动物个体重复结构的形态,数目及其进化,如像脊椎动物 的脊椎骨数目与类别。图左为鸡胚胎发育时期的脊椎组成,Hoxc-6基因 控制7数胸椎的模式,蓝色部分将发育为肋骨。 图右为乌蛇 胚胎,Hoxc6基因的表达调控使乌蛇胸椎数目(蓝色)极度扩展,从头部一直延伸到 尾部。
蛋白 质创 新
功能 域洗 牌
组织血纤蛋白溶酶原激活因子(tissue plasminogen activator,TPA) 是功能域洗牌的一个例子。 TPA基因有4个外显子,最上游的外显 子编码一个指形模块,来自纤连蛋白。第2个TPA外显子指令一个 生长因子结构域,来自表皮生长因子基因。后面2个外显子编 “Kringle”结构,使TPA与血纤维蛋白凝块结合,来自血纤维蛋白 溶原酶基因。Mol Evol 40:331-336,1995
蛋 白 质 体 外 进 化
体外蛋白质进化的设计原理类似于DNA洗牌。利用嵌合 寡聚DNA引物扩放靶基因或同源基因的外显子,随 后将扩放产物混合,变性后复性,再完成自身引物 PCR。最后通过实验筛选重组基因,检测蛋白活性。
Bioscience and Bioengineering,2006,101:449-456
HOX基 因家族 的加倍 与形态
创新
果蝇含有一个与体节分化有关的HOX基因家族。脊椎动物的祖先整个 Hox基因簇连续两次加倍,形成了4个Hox基因簇。哺乳动物4个Hox基 因簇分布在4条不同的染色体上,分别称为Hox-A、Hox-B、Hox-C和 Hox-D。这4个Hox基因簇中,有的基因簇发生了局部的基因重复或缺 失,因此现存哺乳动物总共只有39个Hox基因。
β-球蛋白基因
加倍趋异
哺乳动物和禽类β-球蛋白 基因有着共同的祖先。 原 始β-球蛋白基因( pβ1和 pβ2 )和调控顺序(LCR和 增强子)在经历一次加倍 后,分为两支进化:一支 向真哺乳亚纲和袋鼠发展; 一支成为禽类现存β-球 蛋白基因(p, Ε,pβA和 pβB )的先祖。 PNAS 98:1327-1329, 2001
外显子 洗牌的 分子机
制
-
逆转录介 导的外显 子整合
L1介导的外显子洗牌 a) 在L1的3‘ 端连接反向标记基因neo,当中插入一段内含 子。SD和SA分别为mRNA加工剪接的供体位和受体位。neo基因两端连接异 源启动子(L1pA和SV40pA)和Poly(A)加尾信号。b) 转化HeLa细胞后,反 义neo mRNA的转录物经逆转录后整合到寄主染色体中。c) 当逆转录产物插 入寄主染色体基因的内含子时,该基因转录后产生融合mRNA。d) 融合 mRNA翻译表达Neo蛋白,使转化细胞获得对G418的抗性,在筛选培养基上 可正常生长。Science 283:1530-1534,1999
Hale Waihona Puke 基因 的裂 变与 融合Durrens P等从12种真菌中根据基因组数据收集了1 680个基因融合和裂 变事件。作者发现,基因融合和裂变的数目和基因组大小有关; 从比例看,融合占优势;融合基因趋向于相同的生化功能。作者 认为基因融合和裂变可作为物种进化距离的标尺,比突变更适合 用来判断物种进化的历史。A) 基因裂变事件;B)基因融合事件。 颜色方块为比对的蛋白质结构域,白色是非比对区域。
HOX基因簇的进化
Hox基因家 族在动物的 发育与体节 模式确定中 起着极其重 要的作用。 哺乳动物的 4个Hox基因 簇的来源可 以追溯到原 始的HoxEvx/Meox基 因的加倍, 趋异与缺失, 代表基因进 化的一种主 要方式。
基因重复与适应性进化球蛋白基因
哺乳动物现存球蛋白基因 经历过前后5次加倍与趋 异。原和原球蛋白的 祖先基因在约5亿年前加 倍,在2亿年之后球蛋 白和球蛋白祖先基因又 出现加倍,随之各自发生 分离。现存的肌球蛋白 和球蛋白的功能发生了 趋异,适应了生物进化的 需要: 胎儿期表达的球蛋白可满 足从母体血液吸收氧的功 能,成年表达的球蛋白与 氧结合能力减弱。