最新淋巴细胞发育进展张毓
淋巴细胞信号转导通路匍察

淋巴细胞信号转导通路匍察淋巴细胞是免疫系统中的重要组成部分,负责识别和攻击体内的病原体。
为了实现这一功能,淋巴细胞需要通过一系列复杂的信号转导通路来传递和响应外界的信号刺激。
本文将对淋巴细胞信号转导通路进行匍察,以期更好地理解免疫系统中的免疫应答机制。
淋巴细胞信号转导通路通常被分为两条主要的信号通路:T细胞受体(TCR)信号通路和B细胞受体(BCR)信号通路。
这两条通路在淋巴细胞中起着关键的作用,通过激活一系列蛋白质和转录因子来调节细胞的增殖、分化和功能。
在TCR信号通路中,TCR与特定的抗原结合后,会引发一系列的信号传递过程。
首先,TCR与辅助蛋白共受体CD3结合,形成TCR-CD3复合物。
这一复合物激活下游的蛋白酪氨酸激酶Lck,进而磷酸化免疫受体酪氨酸激酶(ITAMs)位于CD3链上的胞内区域。
磷酸化的ITAMs再进一步结合并激活锌指蛋白ZAP70。
激活的ZAP70通过磷酸化下游信号分子,包括LAT、SOS1和PLCγ1等,最终导致细胞内信号的增强和转导。
与TCR信号通路类似,BCR信号通路也经历一系列的信号传递过程。
当BCR与特定抗原结合后,它会形成一个受体聚集簇。
这种聚集簇激活下游的酪氨酸激酶,包括Src家族激酶Lyn,进而磷酸化免疫受体聚集区域的免疫受体酪氨酸激酶(ITAMs)。
ITAMs的磷酸化激活下游的蛋白质,如Syk和Btk,最终促使细胞内的信号转导。
除了TCR和BCR信号通路外,淋巴细胞还可以通过共刺激分子和辅助细胞来调节其信号传递。
共刺激分子,如CD28和CD40,可以与淋巴细胞表面的相应配体结合,从而激活细胞内的信号转导通路。
辅助细胞,如树突状细胞和辅助T细胞,通过细胞间的接触和分泌的细胞因子来激活和调节淋巴细胞的免疫应答。
淋巴细胞信号转导通路的研究已经取得了显著的进展,并为我们理解和治疗多种免疫相关疾病提供了重要的线索。
例如,选择性抑制特定信号通路的药物已经被广泛应用于肿瘤治疗,通过抑制淋巴细胞的增殖和功能来控制肿瘤的发展。
淋巴细胞发育进展张毓

BCR刺激后,未成熟B细胞中 刺激后,未成熟 细胞中 细胞中PIP2水解和 刺激后 水解和 PKCβ活化不充分 β PKCβ持续活化令未成熟B细胞获得凋亡抗性 β持续活化令未成熟 细胞获得凋亡抗性 PKCβ功能阻断使成熟 细胞对凋亡高度敏感 β功能阻断使成熟B细胞对凋亡高度敏感
c-Rel缺陷鼠来源 细胞受BCR刺激后出现凋亡 缺陷鼠来源B细胞受 刺激后出现凋亡 缺陷鼠来源 细胞受 CD40信号挽救 细胞凋亡 信号挽救B细胞凋亡 信号挽救
Double positive
TCR CD4 CD4
TCR
CD4 SP
CD4 RTE
TSC
DN
DP
TCR CD8
TCR
CD8
β-selection
Positive selection
CD8 SP
CD8 RTE
Negative selection
Functional maturation
胸腺T细胞发育过程及主要事件 胸腺 细胞发育过程及主要事件
Aire and Negative selection
自身免疫性多内分泌腺病-念珠菌病自身免疫性多内分泌腺病-念珠菌病-外胚层营养不良 APECED
慢性皮肤粘膜白色念珠菌病 甲状旁腺功能减退 肾上腺功能不全
Autoimmune Regulator (Aire)
Aire缺陷与自身免疫 缺陷与自身免疫
Aire驱动 驱动TSA在髓质区胸腺 驱动 在髓质区胸腺 上皮细胞中表达 OVA-specific TCR/insulin promoter-driven OVA双转 双转 基因鼠模型
Aire作用机制 作用机制
调控TSA表达 表达 调控 促进mTEC凋亡 促进 凋亡
黎衍敏树突状细胞的研究1

鼓励多学科交叉研究,利用其他学科的先进技术和方法,推动黎 衍敏树突状细胞研究的快速发展。
THANKS
感谢观看
,避免不必要的免疫反应。
04
黎衍敏树突状细胞在疾病中的作 用
在自身免疫性疾病中的作用
辅助T细胞分化
树突状细胞能够通过表达特定的 分子,辅助T细胞分化为Th1或 Th2细胞,影响自身免疫性疾病
的发展。
参与炎症反应
树突状细胞在自身免疫性疾病的 炎症反应中发挥重要作用,它们 可以分泌促炎因子,促进炎症的
分类
根据其发育来源和表型,树突状细胞可分为两类:骨髓来源的髓系DCs和淋巴组 织来源的淋巴系DCs。
树突状细胞的功能
01
02
03
抗原呈递
树突状细胞能够摄取并加 工处理抗原,将其呈递给 T细胞,激活适应性免疫 应答。
免疫调节
树突状细胞能够分泌多种 细胞因子和炎症介质,调 节免疫应答的强度和方向 。
诱导免疫耐受
黎衍敏树突状细胞具有抗原提呈作用,能够摄取并加工处理抗原,将抗原信息暴露在细胞表面,供T 细胞识别。
详细描述
在免疫应答中,树突状细胞扮演着关键角色,它们能够高效地摄取和加工处理抗原,将抗原信息暴露 在细胞表面,从而激活T细胞。这种抗原提呈作用是树突状细胞的核心功能之一,有助于触发适应性 免疫应答。
免疫调节作用
其生物学特性。
阐明细胞命运的决定机制
03
解析黎衍敏树突状细胞命运决定的分子机制,为干预其功能提
供理论依据。
探索黎衍敏树突状细胞在疾病治疗中的应用
1 2
开发新的治疗策略
基于对黎衍敏树突状细胞的深入理解,开发新的 治疗策略,为疾病的治疗提供新的思路。
陕西医学杂志

陕西医学杂志期刊征稿:陕西医学陕西医学期刊级别:统计源期刊主管单位:陕西省卫生厅主办单位:陕西省中医药研究院出版地方:陕西快捷分类:医学国际刊号:1000-7377国内刊号:61-1104/R邮发代号:52-40创刊时间:1972陕西医学杂志简介期刊之家医学核心学术期刊杂志社结成了学术联盟,如果您有医学学术论文需要发表到统计源期刊(中国科技核心期刊)、中文核心期刊(北大核心)、南大核心(cssci)或SCI检索收录期刊等需求的作者请及时与我们联系,进行学术期刊选刊与时间安排等相关事务联系杨老师微信LunwenFz《陕西医学》(月刊)创刊于1972年,由陕西省中医药研究院主办。
本刊为综合性医药卫生学术刊物。
本刊主要报道本地区及全国的医学科研成果,疾病防治经验,新技术应用总结等;介绍国内外医学领域的新理论、新技术、新经验、新动态,使广大读者从中获得新的知识和技能,提高医学临床与基础理论水平。
读者对象为广大医务工作者。
《陕西医学》杂志坚持“科学性、先进性、实用性和服务性”的办刊方针;以“传递医疗科技资讯、报道医生临床进展、展示医生科研成果、交流医生工作经验”为宗旨。
杂志所刊发的论文理论与实践相结合,是全国各级医院、妇幼保健院、社区卫生服务中心、乡镇卫生院、门诊部、诊所等医疗机构广大临床医生和乡村医生及医学院校师生工作、学习的参考和交流的园地。
陕西医学栏目设置论著临床报道、综述讲座、药物与临床、临床检验、基层医学园地、临床护理、短篇报道、经验交流陕西医学杂志收录/荣誉CA 化学文摘(美)万方收录(中)上海图书馆馆藏国家图书馆馆藏知网收录(中)统计源核心期刊(中国科技论文核心期刊)维普收录(中)中国学术期刊(光盘版)全文收录期刊中国期刊全文数据库(CJFD)中国核心期刊遴选数据库中国科技期刊核心期刊全国中文核心期刊目录基础研究汉防己甲素对人绒毛膜癌BeWo细胞凋亡和细胞周期的影响方静;冀静;安瑞芳;1323-1325核转录因子NF-κB、RET/PTC在桥本甲状腺炎合并甲状腺乳头状癌中的表达及其意义李建英;王娟红;鱼军;牛健;李秀萍;张毓;陈巍;1326-1328+1334法舒地尔对HL-60细胞增殖、凋亡、分化及侵袭作用的影响贾少勋;茹杏丽;王一;1329-1331 miR-9过表达对鼻咽癌细胞迁移和侵袭的影响罗花南;马思敬;易春曦;高滢;侯瑾;闫静;任晓勇;1332-1334芹菜素诱导人乳腺癌MDA-MB-231细胞凋亡的实验研究孙晓力;翟宏军;张心武;马小斌;马双余;1335-1337+1365miR-425调控PDL-1通路调节肾上皮细胞增殖、凋亡作用的实验研究蒋小丽;周慧芳;毛能建;1338-1340去铁胺在小鼠局灶性脑缺血损伤中的保护作用苏永永;吴鹏昌;谢江涛;向毅;王世锋;武兴兴;刘振锋;姜海涛;1340-1342大尺寸骨软骨复合支架修复羊膝关节骨软骨缺损的初步研究边卫国;钟亮;连芩;邱裕生;靳忠民;1343-1345高通量测序平台对卵巢癌基因突变的检测及意义姜向阳;李兴珍;刘文康;吴志新;李帅帅;张恩科;赵清侠;1346-1348microRNA217逆转人宫颈癌细胞顺铂耐药及其作用机制张婷婷;彭慧霞;1348-1350Nrf2及P62在卵巢癌组织中的表达及临床意义李亚玲;王丰艳;李欢;王宝玲;1351-1352+1385 钠尿肽极化液抗心力衰竭作用的实验研究王红;铁茹;常盼;张静;王西辉;吴娟;张学策;朱妙章;于军;1353-1354NVP-BEZ235对人结肠癌细胞SW480体外增殖的抑制作用及其机制探讨马善义;孙兆楼;1355-1357丹参酮ⅡA预处理对心肌缺血再灌注损伤保护作用的研究马春香;孙媛;陈晓斌;张蕾;1358-1359缺血预处理和缺血后处理对脑缺血再灌注损伤保护作用的实验研究种莉;陈丽;张李娜;唐鹏;刘玥;1360-1362下颌全牙列数字印模口内法与口外法的准确性比较朱文忠;张春宝;越野;宋鑫磊;1363-1365 临床研究脑白质疏松症与前庭功能受损的相关性研究闫红静;习旭涛;朱琰;吴银霞;李彦梅;申晓玲;1366-1368BOLD-fMRI联合DTI-FT功能神经导航技术在额顶叶胶质瘤手术中的应用杜昌旺;史罗宁;王茂德;陈伟;姜海涛;王伟;王娟;祁磊;1368-1370蛛网膜下腔阻滞麻醉用于不同年龄老年患者股骨转子间骨折闭合复位内固定术的临床观察顾丽莉;叶雷;1371-1373腹腔镜下子宫肌瘤切除患者术后镇痛的选择焦向阳;周绍文;贺显建;肖志彬;曾甜;1373-1374 电视胸腔镜手术对老年非小细胞肺癌患者术后心肺并发症的影响樊纪丹;王霄霖;杨岷;束余声;1375-1376+1414不同麻醉方式对老年膝关节置换术后患者认知功能及Tau蛋白的影响杨曦;孟双双;张钰;孙伟;1377-1378全胸腔镜肺叶切除术中局部应用洛铂治疗肺癌的临床疗效陈瑜;贺静;1379-1380腹部恶性肿瘤患者术前D-二聚体水平与术后深静脉血栓形成的关系探讨吴为民;李杰;武梦华;1381-1383低剂量胰岛素对非小细胞肺癌化疗的临床研究岳江涛;张瑞星;马继红;刘战飞;1383-1385胰岛素信号转导通路相关蛋白在肺气肿患者肺组织中的表达豆亚伟;王宏涛;田伟;朱建飞;戴云;罗向晖;陈耀华;1386-1388腹腔镜下保留迷走神经的老年胃癌根治术52例分析刘会侠;桂普国;1388-1389早期肠内营养支持对老年胃底贲门癌患者术后营养状况及机体恢复的影响王晓燕;王永军;李腊梅;高亚梅;1390-1391肝癌术后患者卡尔森式疼痛管理理念运用及其对生存质量的影响冯爱芳;董芳芳;郑鑫;鱼晓清;王建民;1392-1393青年乳腺癌21例影像学特征分析张毅力;杜红文;高金海;1394-1396胎膜早破与孕妇沙眼衣原体、解脲支原体感染的关系及其对母婴结局的影响武士敏;张洁;万晓琴;1396-1398三腔尿管在剖宫产术后早期产后出血中的应用广会娟;常晓玫;董俊英;1398-1399西安地区流产绒毛组织72例染色体核型分析李金鸽;王惠仔;吕根芳;1400-1401妊娠期糖尿病孕妇血内脂素的变化及其对新生儿体重的影响康瑛;续靖宁;1402-1404宫腔镜配合药物治疗稽留流产43例临床观察陈蕊;崔明双;李琴琴;杨洋;薛芳芳;张玉荣;1404-1406淋巴细胞主动免疫治疗对封闭抗体阴性复发性流产后患者外周血Th1/Th2/Th17细胞因子表达水平变化史晓艳;陈铭;石玉玲;1407-1409急性冠状动脉综合征患者血清hs-CRP、Hcy、LP-PLA2与血管内超声虚拟组织学斑块特征的相关性研究徐冉;吕健;李大鹏;魏林;1410-1411急性冠脉综合征患者GRACE评分与氯吡格雷抵抗相关性分析朱火兰;杜洁;潘硕;刘仲伟;崔倩卫;张勇;朱舜明;祁杰;1412-1414急性冠脉综合征PCI术后早期恶性室性心律失常的危险因素分析梁娟;1415-1416前庭康复训练在前庭性偏头痛治疗中的应用刘西玲;刘玥;杨丹;王欣;1417-1418DSG-Ⅲ型生物信息红外肝病治疗仪治疗慢性肝病31例疗效观察张欣;田长印;吴凤萍;李梅;翟嵩;王媛;潘国英;邱岭;1419-1421血栓抽吸、经皮冠状动脉介入术联合替罗非班治疗急性ST段抬高型心肌梗死疗效观察丁鹏;雷肖蠢;1421-1423血浆ADMA和血小板膜糖蛋白在急性脑梗死患者中的表达及关系的研究邱伟庆;1423-1425 血清CA125水平与心力衰竭患者预后的相关性分析陈岚;徐进;陈仕明;左宏;1426-1428高频超声联合X线检查对痛风性关节炎患者关节损害的诊断价值张景锋;张春峰;张红艳;李建刚;李奇;1428-1429慢性咳嗽与电子支气管镜下气管-支气管炭末沉积的相关性探讨王惠琴;解岚岚;薛佩妮;李天浩;马建刚;陈方园;1430-1432止血敷料在介入术后股动脉穿刺点止血中的应用蒋锋;王莉;袁婕;李志伟;刘军;1432-1433便携式纤支镜在慢阻肺合并严重呼吸衰竭治疗中的应用价值张治国;1434-1435物理疗法联合妥布霉素地塞米松眼膏治疗睑板腺功能障碍临床观察刘慧峰;王霞;韩双羽;朱江;1436-1437机用单根锉Wave-one、手用镍钛器械ProTaper与不锈钢K锉对下前牙根管的预备及评价付峰勃;杨文静;董锐;王艳丽;1438-1439全视网膜激光光凝术联合药物治疗严重糖尿病视网膜病变疗效观察张斌;孙先勇;1440-1441 磁共振DWI与FLAIR成像早期诊断病毒性脑炎价值比较范跃星;任建政;王峰;张锦;牛素丽;张丽琴;李军;兰海波;1442-1444Keratograph眼表综合分析仪对高度近视患者眼表功能的评估贺贵云;李秋霞;1444-1446彩色多普勒超声在妇科急腹症诊断中的临床应用价值庄碧惠;1447-1448彩色多普勒超声检查在孕中晚期胎儿心脏畸形筛查中的临床价值弓瑜;1449-1450非酒精性脂肪肝与同型半胱氨酸水平及胰岛素抵抗的关系探讨延华;张粉利;高艳琼;王欢;1451-1452两种参数超脉冲二氧化碳点阵激光治疗凹陷痤疮瘢痕疗效比较王泽芳;叶飞伦;杨皓月;唐艳;1453-1454药物与临床司维拉姆治疗维持性透析患者高磷血症有效性和安全性观察牛丹;吕佳;王晓培;邵耀中;路万虹;解立怡;张亚莉;尹爱平;郝亚宁;1455-1457门冬氨酸钾镁注射液联合黄芪注射液在高原地区慢性心力衰竭治疗中的应用原勇;刘俊杰;郑建杰;1457-1458+1481地佐辛与舒芬太尼用于剖宫产术镇痛效果比较苗晓茹;小辉;1459-1460间苯三酚联合缩宫素静滴促宫颈成熟及对产程进展、胎儿血气的影响赵影;张春雨;1461-1462罗哌卡因局部浸润联合地佐辛肌肉注射对多发肋骨骨折患者术后镇痛的影响罗振国;邵冠一;易双强;董补怀;肖莉;张洁;1463-1464+1476盐酸氢吗啡酮复合丙泊酚在无痛宫腔镜检查中的应用彭春晓;冯燕;李秀婷;1465-1467黄芩苷对急进高原青年男性左室功能的影响孙书红;段炜;马兰香;原平利;张磊;1467-1468 不同剂量右美托咪啶行硬膜外麻醉在高龄膝关节置换术中的应用莫凯年;李九会;李文均;李圣君;1469-1470七氟醚联合地佐辛复合麻醉用于小儿腺样体肥大切除术中镇痛的应用研究侯瑞礁;潘方立;1471-1472替格瑞洛在急性ST段抬高型心肌梗死经皮冠状动脉介入治疗中的应用冯燕;郝晓云;赵朝;1473-1474压力雾化复方黄柏液外喷对中毒性表皮松解症皮损愈合的疗效观察周英丽;张亚茹;1475-1476临床检验平均血小板体积检测在评估ST段抬高型心肌梗死合并急性肾损伤风险中的意义程慧;蒋薇;李一;韩敏;1477-1478+1486冠心病病程中联合检测HCY、hS-CRP、LP(a)及D-二聚体的临床意义翟向伟;高宏勇;王燕妮;白玲;雷新军;关宏弟;1479-1481调查研究某三甲医院呼吸科重症监护病房患者医院感染发生的危险因素调查研究冯思芳;张兰;王沙沙;白智庆;汪靖园;1482-1484临床护理综合护理干预对消化系统肿瘤化疗患者胃肠道反应的影响李丽萍;何晴;1485-1486综述FOXJ1基因与肿瘤相关性研究进展王博;薛锋;刘娟妮;陈伟;余明;梁英平;李俊海;1487-1488 致作者1411+1446+1460。
北医基础导师资料
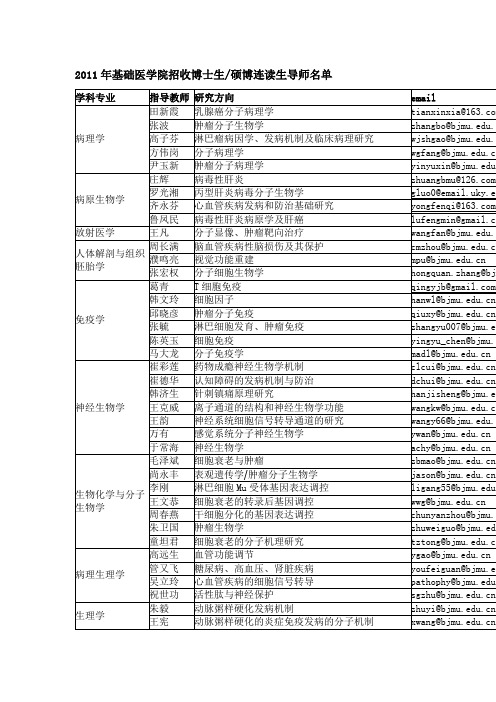
2011年基础医学院招收博士生/硕博连读生导师名单
11年基础医学院招收硕士生导师名单
北京大学医学部基础医学院研究生指导教师
作者:admin文章来源:本站原创点击数:8416 更新时间:2009-2-25
博士生指导教师
生理学刘国庆唐朝枢汪南平王宪徐国恒张炜真朱毅
神经生物学崔彩莲崔德华韩济生万有王克威王韵于常海
细胞生物学李凌松沈丽钟南周柔丽
生物化学与分子生物学贾弘禔李刚毛泽斌尚永丰童坦君王文恭周春燕朱卫国
生物物理学沙印林文宗曜尹长城
放射医学王凡
药理学李学军梁建辉陆林谭焕然杨宝学张永鹤章国良
人体解剖与组织胚胎学濮鸣亮张宏权周长满
免疫学高晓明韩文玲马大龙邱晓彦张毓
病原生物学鲁凤民庄辉
病理学方伟岗顾江尹玉新张波郑杰田新霞高子芬
病理生理学高远生管又飞吴立玲祝世功
中西医结合临床钱瑞琴韩晶岩
硕士生指导教师
生理学黄薇孔炜梅林吴白燕张弘
神经生物学吴鎏桢邢国刚
细胞生物学杜晓娟肖军军张页
生物化学与分子生物学李载权倪菊华张晓伟
生物物理学贺东奇常振战
药理学邓艳萍李卫东时杰
人体解剖与组织胚胎学李英唐军民卫兰徐健张卫光
免疫学陈英玉郭长占王露王应王月丹徐晓军尹艳慧张君赵红珊
病原生物学李杰李彤刘学恩彭宜红王玲朱永红齐永芬
放射医学杨业鹏
病理学宫恩聪刘从容石雪迎钟延丰
病理生理学郑乐民
中西医结合基础王传社。
利用N 糖苷酶F 对单克隆抗体N 糖酶解条件的优化

生物技术进展2021年㊀第11卷㊀第2期㊀214~222CurrentBiotechnology㊀ISSN2095 ̄2341研究论文Articles㊀收稿日期:2021 ̄01 ̄14ꎻ接受日期:2021 ̄01 ̄27㊀基金项目:上海市浦东新区科技发展基金(PKX2019 ̄S09)ꎮ㊀联系方式:张博慧E ̄mail:yaoyaozhd@163.comꎻ∗通信作者许俊彦E ̄mail:junyan.xu@dragonboatbio.com利用N糖苷酶F对单克隆抗体N糖酶解条件的优化张博慧ꎬ㊀贾戴辉ꎬ㊀程倩ꎬ㊀许俊彦∗ꎬ㊀邵喆ꎬ㊀黄应峰宝船生物医药科技(上海)有限公司药物分析部门ꎬ上海201203摘㊀要:为了优化利用N糖苷酶F(PNGaseF)酶解单克隆抗体中N糖的方法ꎬ应用本公司生产的单抗对PNGaseF酶的酶解条件进行优化ꎬ包括缓冲液pH㊁酶种类㊁仪器㊁酶解程度㊁变性缓冲液及酶加入量等ꎬ总结酶解条件对N糖谱结果的影响ꎮ结果显示ꎬ置换缓冲液至1ˑPBS中可以避免某些单抗在低pH酶解时G0F转化为G0F ̄GN并可改善峰型ꎻ快速PNGaseF和加入变性缓冲液能有效提高酶解效率ꎻUPLC和1.7μm粒径色谱柱能提高分离度ꎻ不完全酶解影响N糖含量结果ꎮ研究结果表明ꎬ采用优化的酶解条件可快速㊁有效的酶解单抗上的N糖ꎬ使N糖谱结果准确可靠ꎬ为细胞株筛选和单抗药物质量控制提供有效手段ꎮ关键词:单克隆抗体ꎻN糖ꎻPNGaseF酶DOI:10.19586/j.2095 ̄2341.2021.0005中图分类号:Q819ꎬR97㊀㊀㊀文献标识码:AOptimizationofHydrolysisConditionsofN ̄glycanfromMonoclonalAntibodiesbyPNGaseF㊀ZHANGBohuiꎬJIADaihuiꎬCHENGQianꎬXUJunyan∗ꎬSHAOZheꎬHUANGYingfengDrugAnalysisDepartmentꎬDragonboatBiopharmaceuticalCo.ꎬLtdꎬShanghai201203ꎬChinaAbstract:InordertodevelopandoptimizeamethodofhydrolyzingN ̄glycanfrommonoclonalantibodiesbyusingPNGaseFꎬaseriesofexperimentalconditionsꎬincludingbufferpHꎬenzymetypeꎬinstrumentsꎬdegreeofenzymatichydrolysisꎬdenaturingbuffersolutionandenzymedosagewereoptimizedbyusingseveralmonoclonalantibodies.Resultsshowedthatꎬbufferexchanginginto1ˑPBScouldavoidtheconversionofsomemAbsfromG0FtoG0F ̄GNatlowpHandimprovethepeakshape.RapidPNGaseFanddenaturebuffercouldeffectivelyimprovetheefficiencyofenzymatichydrolysis.CombinationofUPLCandchromatographiccolumnwith1.7μmparticlesizecouldimprovetheresolution.IncompleteenzymatichydrolysiscouldaffecttheresultsofN ̄glycancontent.ResultsindicatedthattheoptimizedhydrolyzingconditionsofPNGaseFcanquicklyandeffectivelyhydrolyzetheN ̄glycanfromthemonoclonalantibodyꎬwhichcanprovideaneffectivemethodforcontrollingN ̄glycancontentincelllinescreeningandqualitycontrolofmonoclonalantibodydrugs.Keywords:monoclonalantibodyꎻN ̄glycanꎻpeptideNglycosidaseF(PNGaseF)㊀㊀N糖基化修饰是单克隆抗体药物普遍存在的翻译后修饰现象ꎮ目前研究表明ꎬ糖基化修饰与单克隆抗体药物的功能㊁药物代谢㊁免疫原性等关系密切[1 ̄3]ꎬ如核心岩藻糖缺失能显著增强ADCC效应[4]ꎬ半乳糖能够增强CDC活性[5]ꎻ末端N ̄乙酰葡萄糖影响抗体药物半衰期[6 ̄7]ꎻ唾液酸化糖型具有抗炎症功能[8]ꎻα(1 ̄3)半乳糖和NGNA型唾液酸易引起免疫原性[9]等ꎮN糖对抗体结构也具有重要作用ꎬ能稳定CH2结构域ꎬ去糖抗体稳定性会变差ꎬ更易发生去折叠和聚集[10]ꎮN糖类型和含量受细胞株㊁培养条件㊁纯化和储存[11 ̄13]等过程的影响ꎬ因此在单抗药物细胞筛选㊁工艺优化㊁纯化㊁产品放行和稳定性考察中控制和监测N糖含量ꎬ保证单抗药物安全性㊁有效性等尤为重要ꎮ目前检测N糖的方法主要有亲水色谱-荧光法[14]㊁CE ̄LIF法[15 ̄16]和质谱法[17 ̄18]. All Rights Reserved.等ꎮ其中亲水色谱-荧光法是目前应用最为广泛的方法ꎬ该方法通常需要用糖苷酶水解糖蛋白上的N糖链ꎬ再用标记试剂进行衍生化ꎬ采用亲水色谱柱分离ꎬ荧光检测器检测ꎮ由于PNGaseF几乎能水解所有哺乳动物细胞产生的N糖ꎬ因此得到广泛应用[19]ꎮ传统的N糖谱检测方法一般耗时较长ꎬ目前快速N糖样品制备的试剂盒已经商品化ꎬ大幅提高了N糖的检测效率ꎬ但是通常价格昂贵ꎮ而应用快速PNGaseF酶与传统制备过程相组合成为提高检测效率和降低检测成本的一种选择ꎮ本研究基于亲水色谱-荧光法ꎬ针对PNGaseF水解条件进行了优化ꎬ发现和解决了酶解过程中易对结果产生影响的一些因素ꎬ建立了高效准确的N糖检测方法ꎬ以期为N糖的检测研究提供参考ꎮ1㊀材料与方法1.1㊀实验材料1.1.1㊀供试品㊀本研究所用mAb1㊁mAb2和mAb2 ̄F均为宝船生物医药科技(上海)有限公司生产的IgG1型单克隆抗体ꎮ1.1.2㊀试剂和耗材㊀N糖苷酶F(PNGaseF)㊁快速PNGaseF酶(rapidPNGaseF)及变性缓冲液(5ˑ)均购自NewEnglandBioLabs公司ꎻ无水乙醇㊁甲酸铵㊁乙酸㊁二甲基亚砜(DMSO)㊁2 ̄氨基苯甲酰胺(2 ̄AB)㊁氰基硼氢化钠㊁β ̄巯基乙醇等均购自Sigma公司ꎻ甲酸和乙腈购自Fisher公司ꎻPBSBuffer(1ˑ)购自上海生工ꎻ10kD超滤离心管购自Merck公司ꎻ100mmol L-1Tris ̄HCl(含1%SDSꎬpH9.0)溶液和非涂层-熔融石英毛细管购自Beckman公司ꎻ纯化柱(GlycoCleanTMSCartridges)购自Prozyme公司ꎻ色谱柱ACQUITYUPLCGlycanBEHAmideColumn(1.7μmꎬ2.1mmˑ150mm)购自Waters公司ꎻ色谱柱AdvanceBioGlycanMappingColumn(2.7μmꎬ4.6mmˑ150mm)购自Agilent公司ꎮ1.1.3㊀主要仪器设备㊀高效液相系统(HPLCꎬ1260)购自Agilent公司ꎻ超高效液相系统(UPLCꎬH ̄ClassPlus)购自美国Waters公司ꎻ液质联用系统(LC ̄MSꎬQExactive)和数据处理软件(BiopharmaFinder)均购自美国Thermo公司ꎻ毛细管电泳仪(CEꎬPA800Plus)购自Beckman公司ꎻ真空离心浓缩干燥仪(RVC2 ̄25)购自德国Christ公司ꎮ1.2㊀实验方法1.2.1㊀蛋白沉淀和N糖衍生化㊀向酶解后的样品中加入3倍体积的冰乙醇ꎬ-15~-25ħ放置约1hꎬ高速离心10minꎬ取上清干燥ꎮ加入10μL2 ̄AB衍生化试剂ꎬ65ħ避光孵育3hꎮ用纯化柱纯化后干燥ꎬ50%乙腈复溶ꎬ准备上机分析ꎮ1.2.2㊀HPLC色谱条件㊀仪器采用高效液相系统(Agilentꎬ1260)ꎻ色谱柱采用AdvanceBioGlycanMappingColumnꎻ流动相A为100mmol L-1甲酸铵(pH4.5)ꎬ流动相B为100%乙腈ꎻ进样量2μLꎻ流速0.5mL min-1ꎻ荧光检测器激发波长260nmꎬ发射波长430nmꎻ色谱柱温度55ħꎻ梯度洗脱ꎮ1.2.3㊀UPLC色谱条件㊀仪器采用超高效液相系统(WatersꎬH ̄classplus)ꎬ色谱柱采用ACQUITYUPLCGlycanBEHAmideColumnꎮ流动相A为100mmol L-1甲酸铵(pH4.5)ꎬ流动相B为100%乙腈ꎬ进样量2μLꎬ流速0.5mL min-1ꎬ荧光检测器激发波长330nmꎬ发射波长420nmꎬ色谱柱温度60ħꎬ梯度洗脱ꎮ1.2.4㊀质谱参数㊀应用液质联用系统(LC ̄MSꎬQExactive)的HESI正离子模式(HESI+)ꎬ离子源的温度和电压分别为320ħ和3.8kVꎬ离子传输管温度为200ħꎬ其他仪器参数设置均经过优化以获得最佳信号响应ꎻ母离子扫描范围700~3000m z-1ꎬ分离度15000ꎬ分析时长60minꎻ数据处理软件采用BiopharmaFinderꎮ1.2.5㊀还原毛细管凝胶电泳(RCE ̄SDS)㊀将蛋白沉淀用100mmol L-1Tris ̄HCl(含1%SDSꎬpH9.0)溶液复溶ꎬ加入β ̄巯基乙醇还原ꎮ仪器采用毛细管电泳仪(BeckmanꎬPA800plus)ꎬ毛细管采用非涂层-熔融石英毛细管ꎬPDA检测器ꎬ波长220nmꎬ电动进样(-5kV)20sꎬ电压分离(-15kV)40minꎮ2㊀结果与分析2.1㊀缓冲液pH对结果的影响分别将mAb1和mAb2样品调pH至4㊁5㊁6和7ꎬ取200μg进行PNGaseF酶解24hꎬ检测N糖含量ꎮ实验结果如表1和图1所示ꎬmAb2在不512张博慧ꎬ等:利用N糖苷酶F对单克隆抗体N糖酶解条件的优化. All Rights Reserved.同pH缓冲液酶解的N糖含量高度一致ꎬ说明pH对该样品N糖水解无影响ꎻ而mAb1中N糖含量随pH的变化出现明显差异ꎬ其中缓冲液pH为4时ꎬ峰2(p2)含量出现大幅度升高ꎬ相应的峰3(p3)含量明显下降ꎬpH6和7条件下N糖含量基本一致ꎮ表1㊀mAb1和mAb2在不同pH缓冲液中酶解的N糖含量Table1㊀N ̄glycancontentofmAb1andmAb2hydrolyzedindifferentpHbuffer样品名称pHN糖含量/%p1p2p3p4p5p6p7mAb142.023.457.66.23.41.60.453.74.276.65.14.22.20.463.91.379.84.84.32.20.473.91.080.24.74.42.30.4mAb24/7.446.05.324.18.24.85/7.645.85.423.98.24.86/7.645.75.424.08.24.97/7.445.75.424.08.24.9图1㊀mAb1和mAb2在不同pH缓冲液中酶解的N糖色谱图Fig.1㊀N ̄glycanchromatogramofmAb1andmAb2hydrolyzedindifferentpHbuffer612生物技术进展CurrentBiotechnology. All Rights Reserved.㊀㊀将mAb1不同pH酶解后的蛋白进行还原CE ̄SDS检测ꎬ结果(图2)显示含N糖重链(HC)基本已被酶解成非糖基化重链(NGHC)ꎬ说明不同pH缓冲液下PNGaseF酶解N糖24h后均酶解完全ꎬN糖含量的变化并非不完全酶解造成ꎮ利用液质联用鉴定各N糖的种类ꎬp2和p3分别为G0F ̄GN(A1F)和G0F(A2F)ꎮ通过比较不同时间(1㊁8和24h)酶解结果(表2)ꎬ发现缓冲液pH为4和5时ꎬp2和p3含量的变化随着酶解时间的增加而呈梯度变化ꎬpH4时尤为明显ꎻpH为6和7时ꎬ变化不明显ꎮ因此推测在某些单抗分子上ꎬG0F上的N ̄乙酰氨基葡萄糖在低pH条件下易从N糖链上脱落ꎬ使G0F形成G0F ̄GNꎬ从而引起G0F ̄GN和G0F含量的变化ꎮ因此在进行PNGaseF酶解时ꎬ应避免酶解体系中pH过低ꎮ图2㊀mAb1不同pH酶解蛋白RCE ̄SDS电泳图Fig.2㊀ReducedCE ̄SDSofdeglycoproteinfrommAb1hydrolyzedindifferentpHbuffer表2㊀mAb1在不同pH缓冲液中酶解不同时间的N糖含量Table2㊀N ̄glycancontentofmAb1hydrolyzedindifferentpHbufferatdifferenttimespH时间/hN糖含量/%p1p2p3p4p5p6p7Man4G0F ̄GNG0FMan5G1FaG1FbG2F419.31.070.78.04.51.90.883.47.373.05.84.32.00.4242.023.457.66.23.41.60.4514.50.878.66.04.62.20.483.82.378.55.24.42.30.4243.74.276.65.14.22.20.4614.50.779.15.74.52.20.483.91.080.44.94.42.40.3243.91.379.84.84.32.20.4714.40.779.75.44.52.20.483.90.880.44.84.42.40.3243.91.080.24.74.42.30.4712张博慧ꎬ等:利用N糖苷酶F对单克隆抗体N糖酶解条件的优化. All Rights Reserved.2.2㊀置换缓冲液的选择为避免样品缓冲液pH及成分对PNGaseF酶解的影响ꎬ用超滤离心管将样品换液至合适的溶液中再进行PNGaseF酶解ꎮ本研究选择了1ˑPBS溶液(pH7.4)和超纯水分别置换mAb1和mAb2的样品缓冲液ꎬ以未置换缓冲液(pH7)为对照ꎬPNGaseF酶解24hꎬ进行N糖谱检测ꎮN糖色谱图和含量分别见表3和图3ꎬ结果表明1ˑPBS溶液㊁超纯水和样品缓冲液(pH7)的N糖谱结果一致ꎮ因为蛋白在超纯水中稳定性较差ꎬ所以本研究选用1ˑPBS溶液(pH7.4)置换样品的缓冲液ꎮ表3㊀mAb1和mAb2在样品缓冲液㊁1ˑPBS和超纯水中的N糖含量Table3㊀N ̄glycancontentofmAb1andmAb2hydrolyzedinsamplebufferꎬ1ˑPBSandwater样品名称缓冲液N糖含量/%p1p2p3p4p5p6p7mAb1样品缓冲液(pH7)3.91.080.24.74.42.30.41ˑPBS3.90.880.44.84.42.40.3超纯水3.91.080.44.94.42.40.3mAb2样品缓冲液(pH7)/7.445.75.424.08.24.91ˑPBS/7.645.95.424.08.24.8超纯水/7.745.85.424.08.24.8图3㊀mAb1和mAb2在样品缓冲液㊁1ˑPBS和超纯水中的N糖色谱图Fig.3㊀N ̄glycanprofilesofmAb1andmAb2hydrolyzedinsamplebufferꎬ1ˑPBSandwater2.3㊀不同PNGaseF酶对N糖结果的影响直接取200μg样品ꎬ分别加入快速PNGaseF酶(rapidPNGaseF)和PNGaseF进行酶解ꎬN糖谱如图4所示ꎮ结果显示用rapidPNGaseF进行酶解时ꎬ若选择用UPLC和1.7μm粒径的ACQUITYUPLCGlycanBEHAmideColumn色谱柱ꎬ则由于分离度的提高ꎬmAb2的N糖色谱图(图4B)中G0F和G1Fa出现肩峰ꎬ而HPLC结果812生物技术进展CurrentBiotechnology. All Rights Reserved.A:mAb2未置换缓冲液HPLC检测ꎻB:mAb2未置换缓冲液UPLC检测ꎻC:mAb2置换缓冲液UPLC检测ꎮ图4㊀不同PNGaseF的N糖谱结果Fig.4㊀N ̄glycanprofilesofmAb2hydrolyzedbydifferentPNGaseF(图4A)由于分离度较差ꎬ此峰未得到有效分离ꎮ当将mAb2样品缓冲液置换至1ˑPBS中时(图4C)ꎬ该肩峰消失ꎮ综合2.1~2.3部分的研究结果可知ꎬ将样品置换缓冲液至合适的溶液中再进行PNGaseF酶解ꎬ对改善N糖谱峰形和结果准确度至关重要ꎬ本研究采用了1ˑPBS溶液ꎬ效果良好ꎮ另外ꎬ在置换缓冲液的基础上使用快速PNGaseF能将酶解时间缩短为数十分钟ꎬ有效提高了检测效率ꎮ选择UPLC能有效提高分辨率ꎮ2.4㊀蛋白变性对酶解效率的影响本研究发现ꎬ在非变性条件下利用快速PNGaseF酶解不同单抗所需酶解时间差异明显ꎬ从数分钟至数小时不等ꎮ为了有效提高酶解效率ꎬ选择所需酶解时间最长的mAb2 ̄F置换缓冲液至1ˑPBS中ꎬ加入变性缓冲液(5ˑ)和1μLrapidPNGaseF酶ꎬ50ħ孵育10min后进行N糖谱检测处理ꎬ将沉淀的蛋白进行还原CE ̄SDS检测ꎬ以未加变性缓冲液(5ˑ)样品为对照ꎮ由图5还原CE ̄SDS结果可知ꎬ相同的酶解时间条件下ꎬ蛋白变性后由于高级结构被破坏ꎬN糖充分暴露与酶接触ꎬ所以其酶解效果更好ꎮ2.5㊀酶解程度对N糖结果的影响在非变性条件下加入不同体积的rapidPNGaseF酶对mAb2 ̄F进行N糖谱检测处理ꎬ将沉淀的蛋白进行还原CE ̄SDS检测ꎬ以加入变性缓冲液(5ˑ)和1μLrapidPNGaseF酶的样品为对照ꎬ酶解条件均选择50ħ㊁10minꎮ由图6(左)912张博慧ꎬ等:利用N糖苷酶F对单克隆抗体N糖酶解条件的优化. All Rights Reserved.可知ꎬ在非变性条件下酶解相同时间ꎬ酶量的增加使非糖基化组分增多ꎬ但酶量增加至2μL时酶解效果仍较差ꎬ而变性条件下用1μLrapidPNGaseF酶解ꎬ糖基化组分基本被转换成非糖基化组分ꎮN糖水解的程度不同ꎬN糖含量(表4)明显成梯度变化ꎬ说明PNGaseF对不同类型的N糖水解效率不同ꎬ因此为了获得样品中真实的N糖含量水平ꎬ应将N糖尽量酶解完全ꎮ图5㊀mAb2 ̄F在变性和非变性条件下切糖后还原CE ̄SDS图谱Fig.5㊀RCE ̄SDSprofilesofmAb2 ̄Fdeglycatedinnatureanddenatureconditions表4㊀不同体积rapidPNGaseF酶解mAb2 ̄F的N糖含量Table4㊀N ̄glycancontentofmAb2 ̄FdeglycatedbydifferentrapidPNGaseFamountRapidPNGaseF/μLN糖含量/%G0Man5G1aG1bG20.543.418.418.45.52.61.048.012.920.16.42.92.054.57.921.27.33.11+变性55.27.520.08.03.13㊀讨论N糖检测结果的准确性受多种因素的影响ꎮ在缓冲液pH对PNGaseF酶解的影响研究中发现ꎬ低pH能使某些单抗在酶解过程中G0F逐渐转化为G0F ̄GNꎬ造成检测结果的严重偏差ꎬ这种情况通常出现在ProteinA亲和纯化后的样本ꎬ在低pH洗脱后直接进行酶解ꎬ若先将样品缓冲液pH调至中性或进行换液处理ꎬ能够避免pH的影响ꎮ对照mAb2置换和未置换样品缓冲液酶解结果ꎬ置换缓冲液能有效消除G0F和G1Fa的肩峰ꎬ消除样品缓冲液成分对检测结果的影响ꎮ另外ꎬ本研究对多种单抗的研究发现ꎬ酶解程度不同的N糖检测结果存在明显差异ꎬ说明N糖苷酶F对不同类型的N糖水解效率不同ꎬ这与已有文献报道[20]一致ꎮ因此为了获得样品中真实的N糖含量水平ꎬ应将N糖尽量酶解完全ꎮ酶解程度可采用将蛋白沉淀进行还原CE ̄SDS检测ꎬ观察非糖基化组分含量的变化情况ꎮ在细胞株筛选和工艺优化阶段ꎬ快速㊁准确的N糖检测结果能够有效推进开发进度ꎬ而该阶段022生物技术进展CurrentBiotechnology. All Rights Reserved.图6㊀不同体积rapidPNGaseF酶解mAb2 ̄F的RCE ̄SDS和N糖谱图Fig.6㊀N ̄glycanandRCE ̄SDSprofilesofmAb2 ̄FdeglycatedbydifferentrapidPNGaseFamount巨大的样本量又对成本控制提出更高的要求ꎬ因此建立一种高效㊁准确且低成本的N糖检测方法尤为重要ꎮ本研究确定了较为通用的PNGaseF酶对单克隆抗体的酶解条件ꎬ即为将样品置换缓冲液至1ˑPBSꎬ取适量样品加入变性缓冲液(5ˑ)和1μLRapidPNGaseF酶ꎬ50ħ孵育10minꎬ进行后续除蛋白㊁干燥等处理ꎬ快速PNGaseF酶和变性处理的应用ꎬ将酶解时间大幅缩短ꎬ提高了检测效率ꎻ同时采用传统的UPLC和ACQUITYUPLCGlycanBEHAmideColumn色谱柱ꎬ有效提高了分离度ꎬ并控制了检测成本ꎮ参㊀考㊀文㊀献[1]㊀FRIESSWꎬSEIDLAꎬSOERGELFꎬetal..N ̄glycosylationheterogeneityandtheinfluenceonstructureꎬfunctionandphar ̄macokineticsofmonoclonalantibodiesandFcfusionproteins[J].Eur.J.Pharm.Biopharm.ꎬ2016ꎬ100:94-100. [2]㊀刘晓宇.单克隆抗体糖基化修饰的研究现状和进展[J].中国生物制品学杂志ꎬ2020ꎬ33(2):216-221. [3]㊀LIUL.Antibodyglycosylationanditsimpactonthepharmaco ̄kineticsandpharmacodynamicsofmonoclonalantibodiesandFc ̄fusionproteins[J].J.Pharm.Sci.ꎬ2015ꎬ104(6):1866-1884.[4]㊀SHIELDSRLꎬLAIJꎬKECKRꎬetal..Lackoffucoseonhu ̄manIgG1N ̄linkedoligosaccharideimprovesbindingtohumanFcgammaRIIIandantibody ̄dependentcellulartoxicity[J].J.Biol.Chem.ꎬ2002ꎬ277:26733-26740.[5]㊀BOYDPNꎬLINESACꎬPATELAK.Theeffectofthere ̄movalofsialicacidꎬgalactoseandtotalcarbohydrateonthefunctionalactivityofCampath ̄1H[J].Mol.Immunol.ꎬ1995ꎬ32:1311-1318.[6]㊀JONESAJꎬPAPACDIꎬCHINEHꎬetal..Selectiveclear ̄122张博慧ꎬ等:利用N糖苷酶F对单克隆抗体N糖酶解条件的优化. All Rights Reserved.anceofglycoformsofacomplexglycoproteinpharmaceuticalcausedbyterminalN ̄acetylglucosamineissimilarinhumansandcynomolgusmonkeys[J].Glycobiologyꎬ2007ꎬ17:529-540.[7]㊀KECKRꎬNAYAKNꎬLERNERLꎬetal..CharacterizationofacomplexglycoproteinwhosevariablemetabolicclearanceinhumanisdependentonterminalN ̄acetylglucosaminecontent[J].Biologicalsꎬ2008ꎬ36:49-60.[8]㊀NIMMERJAHNFꎬRAVETCHJV.Theanti ̄inflammatoryac ̄tivityofIgG:theintravenousIgGparadox[J].J.Exp.Med.ꎬ2007ꎬ204:11-15.[9]㊀王冲ꎬ郭怀祖.不同细胞系表达的抗EGFR单抗糖基化结构对比分析[J].生物工程学报ꎬ2017ꎬ33(6):1018-1027. [10]㊀ZHENGKꎬBANTOGCꎬBAYERR.Theimpactofglycosyla ̄tiononmonoclonalantibodyconformationandstability[J].MAbsꎬ2011ꎬ3:568-576.[11]㊀KILDEGAARDHFꎬFANYZꎬSENJWꎬetal..Glycoprofil ̄ingeffectsofmediaadditivesonIgGproducedbyCHOcellsinfed ̄batchbioreactors[J].Biotechnol.Bioengin.ꎬ2016ꎬ113(2):359-366.[12]㊀BENJAMINDꎬNEHAMꎬMICHAELB.Alowredoxpotentialaffectsmonoclonalantibodyassemblyandglycosylationincellculture[J].J.Biotechnol.ꎬ2017ꎬ246:71-80.[13]㊀STEFANFꎬJOERGHꎬHANNS ̄CHRISTIANM.Glycationduringstorageandadministrationofmonoclonalantibodyformu ̄lations[J].Eur.J.Pharm.Biopharm.ꎬ2008ꎬ70:42-50. [14]㊀丛宇婷ꎬ胡良海.单克隆抗体药物糖基化修饰分析研究进展[J].色谱ꎬ2016ꎬ34(12):1186-1191.[15]㊀MELISSAHꎬYANGWꎬRICHARDR.CharacterizationofN ̄linkedglycosylationinamonoclonalantibodyproducedinNS0cellsusingcapillaryelectrophoresiswithlaser ̄inducedfluores ̄cencedetection[J].Pharmaceuticalsꎬ2013ꎬ6:393-406. [16]㊀GENNAROLAꎬSALAS ̄SOLANOO.On ̄lineCE ̄LIF ̄MStechnologyforthedirectcharacterizationofN ̄linkedglycansfromtherapeuticantibodies[J].Analyt.Chem.ꎬ2008ꎬ80:3838-3845.[17]㊀李媛ꎬ邱建华ꎬ陈家琪ꎬ等.液相色谱-质谱技术对利妥昔单抗及其类似药结构表征和相似性的研究[J].国际生物制品学杂志ꎬ2016ꎬ39(3):116-121.[18]㊀CHENXꎬFLYNNGC.AnalysisofN ̄glycansfromrecombinantimmunoglobulinGbyon ̄linereversed ̄phasehigh ̄performanceliquidchromatography/massspectrometry[J].An ̄alyt.Biochem.ꎬ2007ꎬ370:147-161.[19]㊀TARENTINOALꎬGÓMEZCMꎬPLUMMERTHJ.Deglyco ̄sylationofasparagine ̄linkedglycansbypeptide:N ̄glycosidaseF[J].Bio ̄chemistryꎬ1985ꎬ24:4665-4671.[20]㊀HUANGYNꎬRONO.KineticsofN ̄glycanreleasefromhumanimmunoglobulinG(IgG)byPNGaseF:allglycansarenotcreatedequal[J].J.Biomol.Techn.ꎬ2017ꎬ28:150-157.222生物技术进展CurrentBiotechnology. All Rights Reserved.。
【doc】浅谈人体外周血淋巴细胞染色体的制备

浅谈人体外周血淋巴细胞染色体的制备2008年第2期6月出版食品工程F00DENGINEERINC浅谈人体外周血淋巴细胞染色体的制备Onthepreparationofchromosomeofperipheralbloodlymphocyteofhumanbody董烁谢振兴(河南大学医学院,开封475004)DONGShuo'XIEZhen—xing(MedicalSchool,HenanUniversity,Kaifeng475004,China)摘要染色体制备技术是医学遗传学中最常见而重要的一种技术,由于各个实验室实验条件和个人操作手法的差异,细胞培养和染色体标本的制作方法也不尽相同.笔者就人体外周血淋巴细胞染色体的制片过程中容易出现的问题及近几年制作染色体标本总结的经验和实验方法改进进行探讨,并提出了有效的避免方法.关键词染色体;染色体制备技术;外周血;细胞培养AbstractThepreparationofchromosomeisafrequent andanimportanttechniqueinmedicalgenetics.Some problemsexistedinexperimentwereanalysed.Improvingoftheexperiments,experiencesandprecautionsduring thepreparationofchromosomeofperipheralbloodlym—phocyteofhumanbodytheseyearswerethoroughlydis-cussed.Someeffectivemethodshavebeengiveninthispaper?keywordschromosome;preparationofchromo—some;peripheralblood;cellculture染色体是组成细胞核的基本物质,是基因的载体,他由DNA和蛋白质构成,具有储存和传递遗传信息的作用.人类细胞遗传学研究的主要对象是染色体.生物体细胞染色体数目和结构是重要的遗传标志之一,特别是在真核细胞中.因此深入认识染色体的结构和功能,对生物的遗传,变异和进化以及细胞的增殖,个体的发生和生殖过程的平衡控制都具有十分重要的意义.每个物种的细胞都具有一定的数目,形态,大小的染色体特征,称为染色体董烁,女,1976年出生,1999年毕业于河南师范大学,生物教育专业,助教.收稿日期:2008—03—19组型,各种染色体技术已广泛应用于核型,鉴别变异,体细胞杂交分析和绘制基因图等方面,因此在染色体的分析研究中,制备染色体标本无疑是细胞遗传学最基本而重要的技术,而优良的染色体制片是其他技术的先决条件.正常情况下,人外周血中是没有分裂相细胞的,只有在异常隋况下才能发现.外周血淋巴细胞一般隋况下处于增殖期中的GO(GD期,未经培养的外周血中很难找到正在分裂的淋巴细胞.植物血细胞凝集素(PHA)是人类淋巴细胞有丝分裂的刺激剂,在PHA作用下,能使处于GO期的淋巴细胞转化为淋巴母细胞,进而进行有丝分裂.利用PHA这一特性,淋巴细胞经过含有PHA培养液培养,在体外便可获得丰富的含有丝分裂的生长活跃的细胞群体,再用秋水仙素处理,即可获得终止于分裂中期的淋巴细胞, 进而得到所需的人体染色体图片.加之,淋巴细胞遍及全身,并在所有器官组织中不断循环,能反映个体整体水平情况而不象体细胞那样具有局部性.1细胞培养1.1无污染环境培养环境无毒和无菌是保证细胞生存的首要条件,也是实验成功与否的先决条件.收获细胞前的所有步骤都要保持高度的无菌,严防细菌和病毒的污染.当细胞放置于体外培养时,与体内相比细胞丢失了对生物和有毒物的防御能力,一旦被污染或自身代谢物质积累等,可导致细胞死亡.1.2恒定的温度维持培养细胞旺盛生长,必须有恒定适宜的温度.人体细胞培养的标准温度为36.5±0.5oC,偏离这一温度范围,细胞的正常代谢会受到影响,甚至食品工程2008年第2期6月出版死亡.如果温度高于37℃,细胞的生长速度减慢;如果温度高于40℃,细胞受损,超过43℃,则导致细胞死亡.培养箱的温度应严格控制在37土0.5℃, 为了能精确有效的控温,在普通的隔水式恒温培养箱上可加装一个电子控温仪.若温度过高或过低,则会延缓分裂周期,造成收获时细胞分裂相减少.1.3气体环境气体是人体细胞培养生存必需条件之一,所需气体主要有0和CO.CO既是细胞代谢产物,也是细胞生长繁殖所需成分,他在细胞培养中的主要作用在于维持培养基的pH值.大多数细胞的适宜pH为7.2~7.4,培养基的pH值也应调整到7.2~7.4 之间,偏离这一范围对细胞培养将产生有害的影响. 偏酸时细胞发育不良,偏碱时细胞会出现轻度固缩. 细胞培养液pH的调节最常用的为加NaHCO的方法,因为NaHCO,可供CO,但CO易于逸出,故最适用于封闭培养.1.4培养液植物凝集素(PHA)是体外淋巴细胞培养成败的关键,只有在PHA的作用下才能进入有丝分裂, 因此要考虑他的质量和尝试,质量有问题药效会降低,浓度过高可能会导致红细胞凝集,都会造成实验效果差.2操作步骤2.1细胞接种接种就是将注射器中的血液注入到培养瓶中.将注射器针头刺人培养瓶的胶塞向培养液中加入0.4mL~0.5mL血液.细胞接种通常是在无菌室超净工作台内的酒精灯火焰区进行.接种时无菌操作是否规范,直接影响到细胞培养的成败.如接种时如不慎将手接触到培养瓶的瓶塞,或接触了注射器的针头,可能会造成细菌中霉菌的污染.接种时,加入血液量的多少也会对培养结果有影响,血液太少, 细胞稀薄,细胞生长速度减慢;血液太多,会造成培养时细胞生长时营养成分不够,影响细胞生存.接种操作是在酒精灯附近进行,接种时还应注意避免血液遇高温被破坏.操作时手与酒精灯的距离以火焰不烫手为宜,离火焰太近,血细胞可能被破坏,甚至被烧焦成块,肉眼可见呈团块;同时也会造成针头堵塞.出现这种情况,应及时更换针头.如已接种,应更换一瓶培养液.2.2秋水仙素适量的秋水仙素,适当的处理时机和时间,是获得足够数量的,良好细胞分裂相的条件.分裂相的多少和染色体形态及带型处理是否良好均受其影响. 秋水仙素是一种生物碱,干扰细胞中微管组装,抵制细胞纺缍体的形成,能使细胞分裂停止于分裂中期,以积累大量的中期细胞.其浓度范围较宽,可相差几十倍之多,常用质量浓度为8g/mL~0.5g/mL.一般秋水仙素溶液的浓度与处理时间有一定的关系, 如果在培养中,浓度过低或处理的时间过短,则标本中的分裂细胞就少,染色体瘦长;用量过多,浓度过高,虽能获得中期分裂相,但染色体缩短变粗,不利于核型分析和显带.只有浓度适量,才能确保获得中期分裂相,但染色体缩短变粗,不利于核型分析和显带.只有浓度适量,才能确保获得足够多的分裂相和长度收缩适度的染色体.另外,处理时间延长,染色体发生分离,同样使染色体变粗短,也不利于计数和分析.加秋水仙素的时间是收集细胞制备染色体前2h~3h,加入秋水仙素后,使秋水仙素的终质量浓度为0.04g/mL~0.08g/mL培养液.实验前应根据培养液的用量和秋水仙素的浓度计算出加入的剂量.通常采用的加秋水仙素的器具是容量为1mL的注射器,使用时也应注意控制好注射器活塞,避免推动时用力过猛,导致秋水仙素加入量过多.2.3收获细胞和制片收获细胞和制片包括收集细胞,低渗,固定,滴片染色等几个步骤,每一步操作失误都可能造成最后实验的失败,使玻片上观察不到染色体.2.3.1收集细胞此步骤操作不当,容易造成细胞丢失.培养瓶从恒温箱中取出时,大部分细胞都沉淀在瓶子的底部,在将培养瓶中液体吸入到离心管之前,应用吸管将细胞充分吹打均匀,再移人离心管中.离心之后,吸弃上清,也应小心操作,有时由于吸得过猛,将与上清液相接处的淋巴细胞层吸掉了,造成大量的分裂中期相细胞丢失.2.3.2低渗低渗处理是染色体制备中很重要的环节,是获得分散良好的分裂相的关键步骤,低渗时间和低渗温度都关系到制片的成功.低渗过度或不足都会造成染色体形态不良.在低渗处理时期细胞十分娇嫩,并且表面发黏,低渗细胞混匀时,吹打要适宜,避2008年第2期董烁,等:浅谈人体外周血淋巴细胞染色体的制备6月出版27免细胞破碎及粘团,另外不要将细胞吸到吸管上部及离心管上部,避免细胞丢失.通过对比实验认为采用0.075mol,LKC137cI=处理20min一25min较合适,效果较好.用0.075mol,LKC1溶液于处理中期细胞,使细胞核膨胀破裂,染色体分散良好,便于观察计数.当低渗处理时间过长时,细胞膜过早破裂,导致分裂细胞丢失,或染色体丢失;如果低渗处理时间不足时细胞膨胀不够,则染色体分散不好,染色体仍然成团或相互重叠,不利于进行观察,计数,分析.经过低渗处理的细胞因已膨胀,容易过早破裂,造成分裂细胞丢失,所以低渗后操作应特别注意不要用力冲吸.另外低渗还可使红细胞膜破裂,经离心后,血影浮于上清液中被去除.在观察制片结果时,如发现玻片上染色体有的张不开,有的呈现团状,有的数目不够,这是由于低渗时操作不当引起的.低渗不充分,染色体聚在一起;低渗过头,染色体丢失.低渗是否能达到目的,操作时要注意两个方面:一是要使细胞充分与低渗液接触.这就需要在加入低渗液后,要充分将细胞吹打均匀;二是要注意低渗时的温度和时间.2.3.3固定固定技术也是制备良好分散的染色体的重要步骤.低渗处理完后,需加入固定液,固定的目的是对染色体形态进行固定.若染色体分散不良,可适当加大冰乙酸含量,其同时有改善由于低渗处理不够或固定不充分所造成的缺陷,但过量会造成染色体形态变化和影响分带结果.染色体形态不良与加固定液速度有关,加第一次固定液快或过快,可造成飘带样染色体.固定液每次使用必须新鲜配制,否则将会形成酯类,从而影响固定效果.如果固定液不新鲜或甲醇,冰醋酸的质量不佳时,染色体形象模糊,染色体周围有胞浆背景,固定液为分析纯的甲醇和冰醋酸按3:1的体积比配制.预固定和固定时间:加固定液时,应沿管壁慢慢加入并轻轻吹打均匀,预固定和以后的二,三次固定时,固定液加入太快,混匀太快,会使固定作用过强,染色体扭转;固定作用若不足,染色体出现毛刷状.固定液纯度要高,临时配制,固定后再彻底打匀.吹打细胞时,用力要适度,如吹打过重, 会导致核型中染色体的丢失或变形.采用甲醇和冰醋酸按3:1体积比的固定液进行固定,将常用的第一次固定时间5min,30min改为25min,20min,适当延长时间,固定效果较理想.2.3.4滴片滴片是染色体制备中影响染色体形态的关键一步.首先是载玻片要非常干净,否则会影响染色体的分散和分带效果.滴片用的玻片应干净,无脂,无酸才行.其次是滴片的距离,滴加量多少,制片的方式都会影响染色体分散效果.如果冰片冷冻不够,细胞难以贴附而造成丢失.载玻片经过一系列处理擦干后,放置冰箱内,冰冻4h以上,即为冰片.滴片时现用现拿出,否则会融化,操作时采用0℃预冷的冰片滴片.滴片时导致实验失败的原因有以下几方面:①由于细胞悬液太稀,导致滴片时细胞稀少;②载玻片上水太多,导致细胞悬液顺着水流失;③载玻片不洁净,染色体分散不佳;④滴片后,应立即放入7OcI=的烤箱中,以利于细胞的进一步胀开. Giemsa染色液与pH的影响:染色液浓度高,染色时间应缩短,如果染色液浓度增高,染色时间又延长,染色体着色加深,形态结构不清,对裂隙, 染色体断裂的检出可能减少.pH偏碱,染色体着色发蓝,色泽不艳,pH稍酸,染色体呈玫瑰色,色调鲜艳,形态结构清晰,利于统计分析和畸变的检出.烤片是染色体制备中最后一步技术.烤片的温度,时间与染色体形态和分带有关.不宜温度过高和烤片时间过长.细胞培养和染色体制备实验从接种到制片长达76h,在整个实验过程中必须严谨,不能有任何差错.而且细胞培养实验不同于其他实验,步骤多,整体性强,操作要求严格认真,最终实验才会取得成功.总之,随着医学的发展及人们对遗传疾病的逐步认识,染色体制备技术现已是医学遗传学中最常见而重要的一种技术.作为一名技术人员,做出一张高质量的人体外周血淋巴细胞染色体标本是十分重要的,这样才能更好地为临床诊断及优生与遗传咨询工作服务.参考文献[1]左假.医学遗传学[M],北京:人民卫生出版社.2004:110-127.[2]周焕庚.人类染色体[M].北京:科学出版社.1987:85—86.[3]蔡绍京.细胞生物学与医学遗传实验指南[M].上海:第二军医大出版社,2002:47_48.。
掌握各个阶段淋巴细胞的形态特点及镜下识别

多,对于疾病的诊断和治疗具有重要意义。
免疫缺陷病
03
某些免疫缺陷病患者,外周血中成熟淋巴细胞数量可减少,对
于疾病的诊断和鉴别诊断具有一定帮助。
05 活化淋巴细胞与记忆淋巴 细胞
活化淋巴细胞形态特征及功能
形态特征
活化淋巴细胞体积增大,细胞核增大、分裂活跃,细胞质内 RNA和蛋白质合成增加,可见细胞质扩张和线粒体增多等现 象。
相关疾病与诊断意义
相关疾病
幼稚淋巴细胞增多可见于急性淋巴细胞白血病、慢性淋巴细胞白血病等疾病。
诊断意义
幼稚淋巴细胞是白血病细胞的重要组成部分,其形态学特征是白血病诊断和分 型的重要依据之一。同时,幼稚淋巴细胞计数也是判断白血病治疗效果和预后 的重要指标之一。
04 成熟淋巴细胞阶段
成熟淋巴细胞形态特征
细胞大小
细胞质
成熟淋巴细胞大小较均一,直径一般 在6-9μm之间。
细胞质较少,呈淡蓝色,无颗粒或仅 有少量嗜天青颗粒。
细胞核
成熟淋巴细胞的细胞核呈圆形或卵圆 形,染色质呈粗块状,核膜清晰,核 仁消失。
镜下识别方法及技巧
染色方法
常用瑞氏-吉姆萨染色法进行染色, 使得细胞核和细胞质呈现不同的 颜色,便于观察。
功能
活化淋巴细胞具有高度的增殖能力和分化能力,可迅速分裂 并分化为效应细胞和记忆细胞,参与机体的免疫应答。
记忆淋巴细胞形态特征及功能
形态特征
记忆淋巴细胞体积较小,细胞核较大 而细胞质较少,表面标记和细胞内分 子特征与普通淋巴细胞有所不同。
功能
记忆淋巴细胞具有长期存活的能力, 当机体再次接触相同抗原时,可迅速 增殖并分化为大量效应细胞,产生更 快、更强的免疫应答。
观察要点
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
CD4
CD4 SP
CD4 RTE
TCR CD8
TCR CD8
b-selection
Positive selection
CD8 SP
Negative selection
CD8 RTE
Functional maturation
Aire缺陷与自身免疫
Aire驱动TSA在髓质区胸腺 上皮细胞中表达 OVA-specific TCR/insulin promoter-driven OVA双转 基因鼠模型
Notch1。对诱导和维持B系定向至关重要
Id
IL-7/Stat5
E2A
EBF
Pax5
滤泡B细胞和边缘带B细胞分化
FO:IgM+IgDhiCD23+CD21int MZ: IgMhiIgD+CD23-CD21hi
BCR信号强度: Aiolos,负调控,FO增加 CD21,正调控,MZ增加
Notch2信号:MZ B细胞缺失 BAFF:T2,FO B细胞缺失
阴性选择中的一些问题
Anergy的相对贡献? 受体编辑:25%非转基因B细胞
仅发生在骨髓 如何避免有益BCR被清除?
未成熟B与成熟B细胞对BCR交联反应性差异
BCR刺激后,未成熟B细胞中PIP2水解和 PKCb活化不充分 PKCb持续活化令未成熟B细胞获得凋亡抗性 PKCb功能阻断使成熟B细胞对凋亡高度敏感
VL to JL
CLP
Linc-kit+IL-7R+
Prepro-B
B220+CD19-
Pro-B
B220+ CD19+CD43+
Pax5 Bcl-6 Blimp-1 Xbp-1
Pre-B
Immature B
CD43Pre-BCR+
IgM+IgD-
apoptosis
SelfReactive B
Plasma cell
CD19loCD138+
Mature B
IgMloIgDhi CD21intCD23+
T2
IgMhiIgD+ CD21+CD23+
T1
IgMhiIgDCD21-CD23-
B细胞定向分化的转录调控
E2A: 调控多种B系基因表达,缺陷导致B细胞发育阻滞于CLP阶段 EBF1: E2A的主要靶基因,同时受IL-7/Stat5信号调控 Pax5: 激活B系基因,如CD19、Ig-a,同时抑制非B系基因,如
c-Rel缺陷鼠来源B细胞受BCR刺激后出现凋亡 CD40信号挽救B细胞凋亡
c-Myc过表达赋予未成熟B细胞凋亡抗性 c-Myc过表达干扰转基因鼠B细胞阴性选择
未成熟B细胞中,BCR受抗原刺激后不能定位于脂筏。鉴于PIP2主要位于脂筏,BCR定 位缺陷可能直接影响PIP2水解。BCR定位缺陷可能与胞膜中胆固醇含量低有关。
Aire作用机制
调控TSA表达 促进mTEC凋亡
Aire的其它功能?
并非所有TSA的胸腺表达都受Aire调控,如C反应蛋白、谷氨酸脱羧酶67等。 a胞衬蛋白、二硫化物异构酶A2等的表达并不依赖于Aire,但Aire缺陷鼠依然 表现出针对这些抗原的自身免疫应答。
Aire影响mTEC抗原加工提呈 Aire参与SP T细胞发育调控 Aire参与外周免疫耐受诱导
谢谢结束语谢谢大家聆!!!20淋巴细胞发育进展张毓
Development of Thymocytes
Double negative
Double positive
Pre-TCR
TCR-b rearrangement
TCR
CD4 TCR-a rearrangement
CD8
TSC
DN
DP
TCR CD4
Single positive
CD45-gp38-MHC II+EpCAM+细胞诱导T细胞凋亡
胸腺器官生成
cTEC和mTEC来自共同前体
K5-K8+
K5+K8+
K5-K8+ cTEC K5+K8- mTEC
mTEC发育
B淋巴细胞发育
Ig gene rearrangements
E2A EBF1 Pax5 DH to JH
VH to DHJH