第9章 病毒的遗传分析
遗传学第二版课后部分答案专家讲座

a
c
d
eb
第17页
第七章 细菌遗传分析
10. 在接合试验中,有 Hfr a+ b+ str s × F - a - b - str r,已知 a 和 b 基因控制
营养
需要,先将杂交菌株接种在有链霉素完全培养基上生长,然后在不一样
培养基上测试100个菌落,结果以下:① 添加 a 物质,得40个菌落。② 添
遗传学第二版课后部分答案
第13页
第七章 细菌遗传分析
1. 用一个起源于一个不能合成异亮氨酸细菌菌株(ile -)噬菌体转导一 个不能合成甲硫氨酸细菌菌株(met -)。将接适用肉汤培养基稀释 后涂布在补充有异亮氨酸基本培养基上。另取相同量稀释肉汤培养 基涂布在基本培养基上。基本培养基上长出 18 个菌落, 含有异亮氨酸 基本培养基上长出 360 个菌落。计算标准化重组百分比。
绘制染色体图为何不能以
方式表示?
不能。依据绘制染色体图规则, 图距是依据试验得到重组值去掉 百分号后数字来表示。
遗传学第二版课后部分答案
第4页
第五章 连锁遗传分析
6. 粗糙脉孢菌一个菌株(基因型为 HI)与另一个菌株(基因型为 hi)杂 交, 二分之一后代为 HI 型, 另二分之一后代为 hi 型。请解释该结果。
加 b 物质,得 20 个菌落。③ 基本培养基得 10 个菌落。分别写出这 3 种
菌
Hfr a+ b+ str s × F - a - b - str r
落基因型,计算基因 a 和 b 之间重组率是多少?
40
20
10
MM + a + str MM + b + str
a+ b+ str r(10) a+ b+ str r(10) a - b+ strr(30) a + b- strr(10)
遗传学复习资料

遗传学复习资料遗传学复习资料第⼀章绪论1、遗传:亲代与⼦代之间同⼀性状相似的现象。
2、变异:亲代与⼦代、⼦代与⼦代之间出现性状差异的现象。
3、遗传学模式⽣物——果蝇①只有野⽣型基因存在时,果蝇才长出红眼,该基因突变后,不再长出红眼。
②野⽣型发⽣突变后,出现黄体,则称该突变基因为黄体基因4、孟德尔的豌⾖杂交试验——选择豌⾖的原因:稳定的,可以区分的性状;⾃花(闭花)授粉,没有外界花粉的污染;⼈⼯授粉也能结实。
易栽培,⽣长周期短;种⼦多,便于收集数据;具有许多稳定易区分的性状。
豌⾖花冠各部分结构较⼤,便于操作,易于控制。
成熟后,豌⾖种⼦保留在⾖荚内不会脱落,每粒种⼦的性状不会丢失。
第⼆章、第三章1、减数分裂过程1)减数分裂:是在配⼦形成过程中进⾏的⼀种特殊的有丝分裂。
包括两次连续的核分裂⽽染⾊体只复制⼀次,每个⼦细胞核中只有单倍数的染⾊体的细胞分裂形式。
2)过程:①减数分裂Ⅰ(最复杂最长)A、前期Ⅰ:细线期——出现姐妹染⾊单体,但染⾊质浓缩为细长线状,看不出染⾊体的双重性,核仁依然存在。
在细线期和整个的前期中染⾊体持续地浓缩。
偶线期——同源染⾊体开始联会,出现联会复合体。
(联会复合体=四联体=⼆价体)。
粗线期——染⾊体完全联会,联会配对完毕,缩短变粗,但核仁仍存在。
⼀对配对的同源染⾊体称⼆价体或四联体。
⾮姐妹染⾊单体间可能发⽣交换。
双线期——染⾊体继续变短变粗,双价体中的两条同源染⾊体彼此分开。
在⾮姐妹染⾊单体间可见交叉结构,交叉结构的出现是发⽣过交换的有形结果。
交叉数⽬逐渐减少,在着丝粒两侧的交叉向两端移动,这种现象称为交叉端化。
终变期——染⾊体进⼀步收缩变粗变短,便于分裂移动,分裂进⼊中期。
B、中期Ⅰ:核仁、核膜消失,各个双价体排列在⾚道板上,着丝粒分居于⾚道板的两侧,附着在纺缍丝上,⽽有丝分裂的中期着丝粒位于⾚道板上。
中期I 着丝粒并不分裂。
C、后期Ⅰ:双价体中的同源染⾊体彼此分开,移向两极,但同源染⾊体的各个成员各⾃的着丝粒并不分开。
遗传学(王亚馥-戴灼华主编)课后习题答案

第2章孟德尔式遗传分析: 习题解1 题解a:(1) 他们第一个孩子为无尝味能力的女儿的概率是1/8;(2) 他们第一个孩子为有尝味能力的孩子的概率是3/4;(3) 他们第一个孩子为有尝味能力儿子的概率是3/8。
b:他们的头两个孩子均为品尝者的概率为9/16。
2 题解:已知半乳糖血症是常染色体隐性遗传。
因为甲的哥哥有半乳糖症,甲的父母必然是致病基因携带者,而甲表现正常,所以甲有2/3的可能为杂合体。
乙的外祖母患有半乳糖血症,乙的母亲必为杂合体,乙有1/2的可能为杂合体,二人结婚,每个孩子都有1/12的可能患病。
3 题解:a:该病是常染色体显性遗传病。
因为该系谱具有常显性遗传病的所有特点:(1)患者的双亲之一是患者;(2)患者同胞中约1/2是患者,男女机会相等;(3)表现连代遗传。
b:设致病基因为A,正常基因a,则该家系各成员的可能基因型如图中所示c:1/24 题解a:系谱中各成员基因型见下图b:1/4X1/3X1/4=1/48c:1/48d:3/45题解:将红色、双子房、矮蔓纯合体(RRDDtt)与黄色、单子房、高蔓纯合体(rrddTT)杂交,在F2中只选黄、双、高植株((rrD-T-))。
而且,在F2中至少要选9株表现黄、双高的植株。
分株收获F3的种子。
次年,分株行播种选择无性状分离的株行。
便是所需黄、双、高的纯合体。
6 题解:正常情况:YY褐色(显性);yy黄色(隐性)。
用含银盐饲料饲养:YY褐色→黄色(发生表型模写)因为表型模写是环境条件的影响,是不遗传的。
将该未知基因型的黄色与正常黄色在不用含银盐饲料饲养的条件下,进行杂交,根据子代表型进行判断。
如果子代全是褐色,说明所测黄色果蝇的基因型是YY。
表现黄色是表型模写的结果。
如果子代全为黄色,说明所测黄色果蝇的基因型是yy。
无表型模写。
7 题解: a:设计一个有效方案。
用基因型分别为aaBBCC、AAbbCC、AABBcc的三个纯合体杂交,培育优良纯合体aabbcc。
第三讲 病毒的遗传变异分析

病毒进化的例证
粘液瘤病毒与宿主的共同进化 甲型流感病毒的抗原性转移以及漂移
人类的感染
重组的甲型流感在人群中三次大流行:
1918年西班牙流感
1957年亚洲流感
1968年香港流感
流感期間 Camp Funston 的緊急軍事醫院。
禽流感病毒(Avian influenza virus, AIV)
野生水禽是禽流感病毒的基因库
病毒的变异
甲型流感病毒变异的显著特点是HA和NA发生
遗传性或抗原性漂移(genetic or antigenic drift) 遗传性或抗原性转移(genetic or antigenic shift)
表型的一致性,遗传的动态差异,在变异中 保持稳定。
突变
病毒复制过程中单个或者成段的核苷酸序列发生 替换、缺失、插入等现象。 缺损型干扰(defective interfering, DI)突变 株,突变的特例
缺损型干扰(defective interfering,DI)突变株
这是一种缺失突变的产物。自身不能复制,只有在
“transforming principle” demonstrated with Streptococcus pneumoniae
病毒
第一节 第二节 第三节 第四节
突变 诱变 基因重组 病毒基因产物间的相互作用
准种:由一种母序列和来自该序列的大量突 变基因组所组成的病毒群体。
病毒的遗传与变异
What is a gene?
Stable source of information Ability to replicate accurately Capable of change
(循证医学课件)第9章疾病的病因与危险因素的循证分析与评价(A)

否定了其因果效应。
☻上述情况说明——
病因/危险因素的证据,只有通过人体试验的 RCT最为可靠, 在某些特定条件下求证病因是可以或可能的。
☻如果找不到人体的RCT病因证据,那么应 检索与评价观察性研究的证据。
图 流行病学三角
二、轮状模型 又称车轮模型(wheel model) ☻此模型强调病因来自宿主及环境两方面。
宿主处于环境包围之中,类似轮状。 宿主是遗传作用的内核。 环境包括生物的、理化的和社会的环境 各部分的大小是有伸缩性的,以不同疾病而异。若环 境因素作用大,则图中的外环可变大些。若是遗传病 ,则表示遗传作用的核要变大些,环境“环”相应缩 小些。
(一)不同等级研究设计的证据
1.真正的人体试验性研究——RCT
以健康人体或特定的患者为研究对象,按照试验制订 纳人与排除标准,以及科学的假设要求,选择一定数 量的合格研究对象,并随机暴露于病因/危险因素,然 后观察一定时间、分别测量两组研究对象目标疾病的 发生率,以探求该致病因素发病对危险度RR值为223( 40/3414:1/19525)。
☻又如:
20世纪90年代,北美年轻妇女为美胸,整形医 院开展了硅酮注入乳房成形术,颇受欢迎,于 是接受整形手术的妇女竞达1400万人次之多, 术后有数百人发现局部症状或周围纤维病变, 也有发生关节炎、红斑狼疮、硬皮病患者,对 此,引起了患者向法院诉讼。
WHO确定国际一流的医学检验中心对该 病的病因病原体进行国际协作研究,终 于通过国际广泛合作,弄清了引起该病 的病因是一种冠状病毒变异株。 定名为急性传染性非典型肺炎或急性重 度呼吸道感染综合征,简称“非典”(severe
细菌及病毒的遗传分析h

trp2+ his2+ tyr1+转化trp2- his2- tyr1- 实验 trp2 34 his2 13 tyr1
Hfr菌株在切除F因子时发生错误切除,分离出一个携带F因子和部分宿主染色体基因的遗传因子,这种带有宿主染色体基因的F因子称为F΄因子。
T2噬菌体的基因重组
将两种不同的T2突变体进行杂交,对其杂交子代进行重组分析 杂交方法: 将Ttor和Ttos两种大肠杆菌细胞混合 同时接种高浓度的T2噬菌体的h-r+和h+r-两种突变体,保证绝大多数细菌都被一个以上噬菌体感染 两种不同的噬菌体DNA可能在宿主细胞内进行重组,从而产生非亲本型子代h+r+和h-r-。 亲本型 重组型
F因子在杂交中的行为——接合过程
(三)中断杂交实验作图
中断杂交实验作图
1分钟≈20%的重组值
二、转化
转化(transformation):指某些细菌(或其它生物)能通过其细胞膜摄取周围介质中的DNA片段,并将此外源DNA片段整合到自己染色体组中的过程。 (一)转化的过程 非感受态细胞 外源DNA被洗掉了 转化因子 感受态细胞 外源DNA仍与细胞结合 整合 吸收 整合 供体单链DNA进入受体细胞后与受体染色体的某一部分联会,并进一步置换受体的对应染色体区段的过程。
第十章 细菌及病毒的遗传分析(2h)
1
第一节 细菌和病毒遗传研究的意义
2
第二节 噬菌体的基因重组
3
第三节 细菌基因重组
4
本章要求
5
思考题
繁殖世代所需时间短;
易于管理和进行化学分析;
便于研究基因的作用;
便于研究基因的突变;
遗传物质较简单,便于用作研究基因结构、功能及调控机制的材料。
遗传学_ 细菌和病毒的遗传分析_
1180 + 418 + 685 +107 +11940 +3660
100% = 2390 100% =13% 17990
trp2
tyr
34
his2
13 tyr1
his
40
trp
八、转导(transduction)
⚫ 普遍性转导(Generalized transduction)
转导是以噬菌 体为媒介,将 外源基因携带 入细菌,使受 体细胞发生遗 传重组的方式。
a、b间发生交换
单性状的转化子
a、b间不发生交换
双性状的转化子
七、转化作图的原理
细菌两连锁基因的交换率
=
单性状转化子的数 单性状转化子数+共转化的转化子数
100%
表7-1 枯草芽孢杆菌trp2+ his2+ tyr1+(供体)× trp2- his2- tyr1-(受体)的转化实验 座位转化子类型
噬菌体的遗传分析
一、细菌和病毒的遗传分析
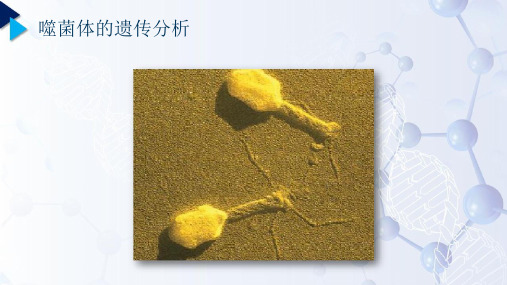
7-1 T4噬菌体的电镜照片
二、病毒对遗传学研究的贡献
1952年 Hershey & Chase的同位素示踪试验
证明T4病毒的遗传物质 是脱氧核糖核酸(DNA) 【1969年诺贝尔奖】
二、病毒对遗传学研究的贡献
1956年Fraemkel Conrat的烟草花叶病毒的重建试验
滑,可致病)
粗糙型R菌株 (无荚膜,菌落粗
糙,不致病)
三、转化现象的发现——Griffth的肺炎双球菌实验
IIR菌株不致病 IIIS菌株致病
灭活的IIIS菌株不致病 灭活的IIIS菌株的某种物 质使IIR菌株发生性状改 变,变成致病的IIIS菌株
遗传学教学大纲

《遗传学》课程教学大纲一.基本信息课程编号:课程名称:遗传学英文名称:genetics课程性质: 专业必修课总学时:54学分: 3适用对象: 生物科学专业先修课程:高等数学植物学动物学生物化学二.编写说明(一)课程的性质遗传学是生物学所有专业的一门专业基础课程,是研究生物遗传和变异的科学,研究内容包括基因的结构与功能、基因从亲代传递到子代过程中的遗传与变异。
随着现代生物科学的发展,遗传学已成为21世纪生命科学领域发展最为迅速的学科之一,是生命科学各门学科的核心,它的分支几乎扩展到生物学的各个研究领域。
本课程的任务是全面系统地讲授遗传学的基本原理和遗传学分析的基本方法,同时介绍现代遗传学发展的最新成就,使学生对遗传物质的本质、遗传物质的传递、遗传物质的变异等基本规律有比较全面的、系统的认识,并能应用其基本原理分析遗传学数据,解释遗传学现象,同时对遗传信息的表达与调控有一个较为全面和深入的了解。
(二)课程教学目标基本要求1.系统了解遗传与变异的规律,分子基础及应用。
2.掌握遗传学的基本实验方法和技能、技巧,并在科学态度、独立工作能力方面获得初步的训练。
3.能够利用所学知识,说明和解决实践中有关遗传学的一般问题。
4.能够根据具体要求和目的查阅相关文献。
5.能够胜任中学生生物课中有关遗传学的讲授、实验和课外活动等教学工作。
(三)课程的重点和难点第二章第二节孟德尔的分离规律和独立分配规律;第二章第二节基因与环境的关系;第三章第二节连锁遗传和性连锁;第五章第二节病毒的遗传;第六章第三节细菌的遗传;第十章第三节细胞质遗传;第十一章第二节染色体结构和数目变异;第十二章第一节基因突变;第十三章第二、三节群体遗传与进化;第十四章第二节数量遗传。
(四)课程教学方法与手段以理论讲授为主,配合实践性教学、实验教学、多媒体教学、学生作业、撰写论文、自学等方法进行学习。
(五)实践环节1.名称洋葱根尖有丝分裂染色体标本制备及观察;植物多倍体人工诱导;大葱花粉母细胞减数分裂标本制备及观察;显微摄影;染色体组型分析;果蝇唾腺染色体制片观察;质粒DNA的提取;蚕豆根尖微核检测技术;遗传学实验的计算机模拟;人群中PTC味盲基因频率的分析;真核基因组DNA的快速提取;染色体分带技术。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
如果编码多肽链中某一氨基酸 的密码子突变为终止密码子,多肽 链合成到此便会中断,从而使多肽 链变短而失去活性。这种突变称为 无义突变(nonsense mutation)。 各种无义突变都是条件致死突变, 因为有相应的无义抑制基因(su+) 。
这些sus突变型之所以在带有相 应的抑制基因宿主中可产生后代, 是因为翻译过程中,在终止密码子 处插入一个特定的氨基酸,防止在 终止密码子位置上提前终止。如琥 珀突变就有许多种抑制基因,各插 入某个氨基酸以防止提前终止(见 表2)。
表1 不同宿主菌中sus突变噬菌体的表型
噬菌体基因型
野生型 sus amb sus och sus op
宿主菌基因型
su+ su+ amb + + su+ och + + + su+ op + +
注:“+”:产生子代,“-”:不产生子代
根据在带有专一性抑制基因的宿主中 的非致死性,可以将sus突变分为3类:琥 珀突变(amber,amb)、赭石突变( ochre,och)和乳白突变(opal,op), 它们相应的密码子为UAG、UAA和UGA。这 些终止密码子不编码任何氨基酸,称为无 义密码子(nonsense codon),是蛋白质 合成的终止信号。
不同速溶性噬菌体的突变在表型上 不同,可分别写成ra-、rb-、rc-等,用 h+rx-×h-rx+获得的试验结果如下:
表3 h+rx-×h-rx+噬菌斑数目及重组值 每种基因型的%
杂交组合 h+ra-×h-ra+ h+rb-×h-rb+
hr
34 32
+ -
hr
42 56
- +
hr
12
+ +
hr
12
Benzer的重组测验
可用以下公式计算相邻不同位点的遗传距离:
2 rII重组噬菌体数 Rf 总噬菌体数
由于在E.coli B平板感染后得到的噬菌斑 数为群体总数,在E.coli K12 (λ )的噬菌斑 为基因内重组所得到的野生型的噬菌体数,所 以以上公式可以表示为:
2 在E.coliK( )上生长的噬菌斑数 Rf 在E.coliB上生长的噬菌斑数
二、基因内互补
通常同一顺反子内两个突变是不能互补的 ,但是也有一些例外。这种例外发生于同一基 因内两个不同位点突变致使同一种多肽转变成 两条分别在不同位点上发生变异的多肽链,而 后将这两条多肽构成双重杂合子,这两者配合 起来,有可能表现出不同程度酶活性部位的恢 复,这种现象称为基因内互补( intragenic/intracistronic complementation),又称等位(基因)互补 (allelies complementation)。
- -
重组值 24%
5.9
6.4 12.30%
h+rc-×h-rc+
39
59
0.7
0.9
1.60%
每一杂交得到一连锁图:
ra
24.0
h
12.3
rb
h
1.6
rc
h
三种不同的重组值,表示三个r基因的 座位是不同的,所以有4种可能的连锁图: ra rb rc h
ra
ra ra
rb
h rc
rc h h rc rb rb
确定四个基因的排列顺序时,首先只 考虑rb,rc和h,进行rc-rb+×rc+rb-的杂 交,得到的重组值与rb-h间的距离进行比 较,若rc-rb>12.3=rb-h,则h位于中间, 即排列顺序为rc-h-rb。 剩下的是ra在h的哪一边?答案是rarc-h-rb和rc-h-rb-ra都正确,因为T2噬菌体 的连锁图也是环状的。
h
rc
rb
图
ra T2的连锁图,只注明4个基因
三、λ 噬菌体的三点测交与作图 Kaiser在1955年最早进行了λ 噬菌体 的重组作图试验。他用紫外线照射处理得 到5个λ 噬菌体的突变系:
s 系:产生小噬菌斑 mi系:产生微小噬菌斑 c 系:产生完全清亮的噬菌斑 co1系:产生除中央一个环之外其余部分都 清亮的噬菌斑。 co2系:产生比co1更浓密的中央环噬菌斑。
表现型 +++ s co1 mi s++ + co1 mi s co1 + ++ mi 数目 975 924 30 32 61 51 占总数比例/% 90.82 2.97 5.35 0.86 100 √ 3.83 √ √ √ 6.21 8.32 √ √ 重组值/% s-co1 co1-mi s-mi
2、噬菌斑形态和宿主范围的突变型
噬菌斑形态突变型:突变后有的是由于侵 染宿主细胞后溶菌速度的快慢而形成大小不同 的噬菌斑(plaque)。另一些则是由于被感染 细菌全部或是部分被杀死而形成清晰或混浊的 噬菌斑。 宿主范围的突变型:噬菌体感染细菌时, 首先吸附于细胞表面的专一受体上,这是由受 体的基因控制的,如果受体发生改变,有可能 使噬菌体不能附着,从而该噬菌体的宿主范围 就缩小,另外,噬菌体突变也可以扩大寄生范 围。
Benzer的互补试验: ①用两个不 同的rⅡ突变型 rⅡA和rⅡB同时 感染E.coli K12 (λ),结果可以 互相弥补对方的 缺陷,共同在菌 体内增殖,引起 溶菌,释放原来 的两个突变型。
②用rⅡA中 的两个突变型 ,共感染或用 rⅡB中的两个 突变型共感染 E.coli K12 (λ )菌株,都不 能互补,不产 生溶菌现象。
s + mi
+ co1 + 合计
5
13 2091
从双交换的类型可知,这3个基因的 次序就是s-co1-mi。s与co1的图距应为 3.83cM;co1 与mi之间则为6.21cM;因为 有双交换的存在,s与mi之间的图距则为 8.32+2×0.86=10.04(cM)。 3个基因的遗传图: s co1 mi
② 噬菌体的基因重组发生在噬菌体的DNA 复制以后,因此从单个混合感染的细菌中可以 得到亲代噬菌体和重组噬菌体; ③ 噬菌体不同基因型之间可以发生多次 交换; ④ 在噬菌体中基因重组率可以随着宿主 细胞裂解时间的延长而增加。 因此噬菌体杂交时,应该注意: ① 控制每种亲代噬菌体基因型的投放量。 ② 控制允许发生复制和重组的时间。
第三节
噬菌体突变型的重组测验
一、Benzer的重组测验与基因的精细 结构分析 二、T2 突变型的两点测交与作图 三、λ 噬菌体的基因重组与作图
一、Benzer的重组测验与基因的精细 结构分析
1955年,Benzer :噬菌体遗传区的精细结构 1、用T4的两种rⅡ突变体共感染E.coli B; 2、形成噬菌斑后收集溶菌液,将此溶菌液等分 成两份; 4、取一份溶菌液,再感染E.coli B;取另一份 溶菌液感染E.coli K(λ )。 5、观察每份溶菌液所产生的噬菌斑的数目。
Kaiser用s co1 mi×+++杂交,后 代有8种类型。病毒是单倍体,与二倍体 生物不同,亲本组合与重组交换的组合 可直接从后代中反映出来。8个类型中数 目最少的两种类型就是双交换的结果, 频率最高的两种是λ 噬菌体s co1 mi×+++杂交结果及重组值计算
二、病毒的基因组
根据病毒基因组的结构、以及其转录 mRNA、指导蛋白质合成与病毒复制表达 的特点,基本上可以将其分成6种类型: 双链(ds)DNA病毒基因组 单链(ss) DNA病毒基因组 正链(+) RNA病毒基因组 负链(-) RNA病毒基因组 双链(ds) RNA病毒基因组 反转录病毒基因组
3.83 6.21
虽然将真核生物中两点测交和三点 测交的基本原理和方法应用于噬菌体杂 交,但是两者在杂交机制上是完全不同 的: ①噬菌体在杂交中,每个亲代对子代 所提供的遗传贡献取决于感染细菌时每 种亲代噬菌体的相对数量。如基因型A与 基因型B的亲代比=10:1,则产生重组 子代的数量A型常多于B型子代数。
r-:噬菌斑形态突变型,快速溶菌,产生大的有 明显界限的噬菌斑。 r+:噬菌斑形态为野生型,缓慢溶解产生界限不 清的小噬菌斑。 h+:能够溶解E.coliB品系,不能溶解E.coliB/2品系 h-:宿主范围突变型,既能溶解E.coliB品系也能 溶解E.coliB/2品系。
h-r+、 h+r-混合感染E.coli B ,子代噬菌体再感染E.coli B和 E.coli B/2,得到四种噬菌斑(透 明、小;半透明、大;半透明、小 ;透明、大)。子代的基因型分别 为h-r+、h+r-、h+r+、h-r-。
第九章 病毒的遗传分析
重点:顺反子的概念;噬菌体的重 组测验、互补测验和缺失作 图。 难点:噬菌体的互补测验和缺失作 图。
第一节 病毒的形态结构与基因组 一、病毒的形态结构 二、病毒的基因组
一、病毒的形态结构
病毒由蛋白质外壳包裹的DNA或RNA 组成。
病毒的分类 通常按宿主类型病毒分为: 1)噬菌体(phage): 细菌病毒、真菌病毒 2)植物病毒(plant virus): 感染高等植物、藻类等真核生物的病毒 3)动物病毒: 昆虫病毒和脊椎动物病毒
2、温和噬菌体的增殖 其感染周期有两种途径:即裂解 途径和溶源途径,分别称为裂解周 期(lytic cycle)和溶源周期 (lysogenic cycle)。 用紫外线或化学物质如丝裂霉 素C诱导,温和噬菌体可从溶源周期 转变为裂解周期。
诱导
二、噬菌体的突变型
1、条件致死突变型 温度敏感突变(temperature sensitive mutation,ts),野生型噬 菌体能在很大的温度变化范围内感染宿 主并进行繁殖,而热敏感突变型通常在 30℃(许可条件)感染宿主进行繁殖, 但在40~42℃(限制条件)就是致死的 ,不能形成噬菌斑;冷敏感突变型在较 低温度下就致死。