蛋白质翻译-精
蛋白质翻译后修饰

细胞应激反应
在应激条件下,如氧化应激和DNA损伤, 蛋白质翻译后修饰可以调控应激反应相关蛋 白的活性和功能,从而影响细胞的生存和凋
亡。
THANK YOU
泛素化作用
泛素化可以影响靶蛋白的稳定性、定位、活性以及与其他蛋白质的相互作用,从 而调控细胞内的多种生物学过程,如细胞周期、信号转导和自噬等。
泛素化可以标记受损或不需要的蛋白质,引导其被蛋白酶体降解,从而维持细胞 内蛋白质的平衡。
泛素化调控
泛素化过程受到严格的调控,涉及多种酶的协同作用。这些酶包括E1(泛素活化酶)、 E2(泛素结合酶)和E3(泛素连接酶)。
E3酶在泛素化过程中起着关键作用,它能够识别并结合特定的靶蛋白,将泛素分子准 确地连接到靶蛋白上。
此外,去泛素化酶能够逆转泛素化过程,去除已经结合在靶蛋白上的泛素分子,从而对 泛素化进行动态调控。
05
其他翻译后修饰
乙酰化
总结词
乙酰化是一种常见的蛋白质翻译后修饰,通过将乙酰基团连接到蛋白质的特定氨基酸残基上,可以调节蛋白质的 活性和功能。
翻译后修饰可以影响蛋白质的稳定性 ,通过增加或减少蛋白质的降解速率 ,从而影响细胞内蛋白质的水平和功 能。
蛋白质降解
某些翻译后修饰,如泛素化,可以标 记蛋白质进行降解,通过蛋白酶体途 径降解蛋白质,维持细胞内蛋白质的 动态平衡。
蛋白质功能调控
酶活性调节
亚细胞定位
许多蛋白质在翻译后被修饰以改变其酶活性, 例如,磷酸化可以激活或抑制酶的活性,从 而调控代谢过程和信号转导。
03
疾病与磷酸化
许多人类疾病与蛋白质磷酸化的异常有关。例如,一些癌症和神经退行
性疾病的发生与特定蛋白质的异常磷酸化有关。因此,对蛋白质磷酸化
分子生物学--蛋白质的翻译课件

Initiation requires factors and free subunits
(2)细菌中有三种起始因子 IF-3:稳定30S亚基;辅助 30S亚基与mRNA上起始点特 异性结合; IF-1:与30S亚基结合在A位, 阻止氨酰-tRNA进入;阻止 30S与50S亚基结合。 IF-2:结合特定起始因子 tRNA,控制它进入核糖体; 有核糖体依赖GTP酶活性;
5.1.2 氨酰-tRNA合成酶引入的两种错误
◆蛋白质合成真实性主要决定于:
tRNA能否把正确的氨基酸放到新生多肽链的 正确位置。 ◆氨酰-tRNA合成酶会引入两种错误:
一种是将错误的氨基酸加在正确的tRNA上; 另一种是将正确的氨基酸加在错误的tRNA上。 前者现错的可能性更大。
Error rates differ at each stage of gene expression
Eplroontegiantibo5yn.2tRr.ai3bno蛋ssfoe白rmfer质omm合ovpee成spta的idloyn三l-gtR个mNRA阶NtAo段,aem简xitne介onadcinygl-tRNA
Termination Polypeptide chain is released from tRNA, and ribosome dissociates from mRNA
I: 次黄嘌呤
1.4.4 读码的连续性
生物合成过程中,mRNA的编码方向是 5`→3`,从N端向C端延伸肽链。一条肽链 合成起始后,密码子按3个一框读下去不重 叠也不跳格,直到终止。
2 蛋白质合成中使用的RNA
概述 2.1 mRNA 2.2 tRNA 2.3 rRNA
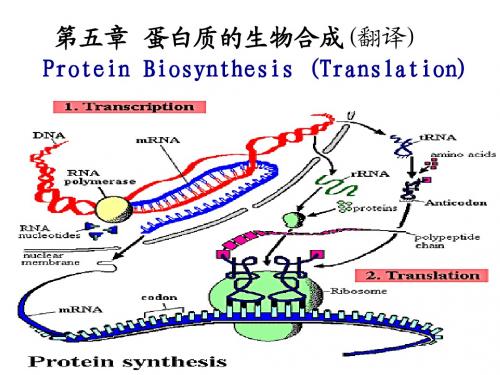
第四章蛋白质的翻译

色氨酸-tRNA的分离
用20种AA-tRNA做20组同样的实验, 每组都含20种AA-tRNA和各种三核苷 酸,但只有一种氨基酸用14C标记,看 哪一种AA-tRNA被留在滤膜上,进一 步分析这一组的模板是哪个三核苷酸, 从模板三核苷酸与氨基酸的关系可测 知该氨基酸的密码子。例如,模板是 UUU时,Phe-tRNA结合于核糖体上, 可知UUU是Phe的密码子。
摆动假说:
在密码子与反密码子的配对中,前两对严格遵守碱基配对原则,第三 对碱基有一定的自由度,可以“摆动”,因而使某些tRNA可以识别1个 以上的密码子;
如果有几个密码子同时编码一个氨基酸,凡是第一、二位碱基不同的 密码子都对应于各自独立的tRNA。原核有30-45种tRNA,真核有50种 tRNA。 I: Inosine is formed by deamination of adenosine after tRNA synthesis
4.2 tRNA
tRNA在蛋白质合成中处于关键地位,它不但为每个三 联密码子翻译成氨基酸提供了接合体,还为准确无误地将 所需氨基酸运送到核糖体上提供了运送载体,所以,它又 被称为第二遗传密码。
tRNA参与多种反应,并与多种蛋白质和核酸相互识 别 ,这就决定了它们在结构上存在大量的共性。
4.2.1 tRNA的结构 1、tRNA的二级结构
1954年科学家对破译密码首先提出了设想: A. 若一种碱基对应与一种氨基酸,那么只可能产生4种氨基酸; B. 若2个碱基编码一种氨基酸的话,4种碱基共有42=16种不同的排列
组合; C. 3个碱基编码一种氨基酸,经排列组合可产生43=64种不同形式; D. 若是四联密码,就会产生44=256种排列组合。
贮 存 在 DNA 上 的 遗 传 信 息 通 过 mRNA 传 递 到 蛋 白 质 上 , mRNA与蛋白质之间的联系是通过遗传密码的破译来实现的。 mRNA上每3个核苷酸翻译成蛋白质多肽链上的一个氨基酸, 这3个核苷酸就称为密码,也叫三联子密码。
9. 蛋白质翻译(1)

摇摆的原因(摇摆假说):
一般地,同义密码子的第1、2位是保守的,而第3位 则是可变的,意味着该可变位点的配对具有一定的灵活 性。
tRNA的反密码子在反密码环上呈弧状排列,与密码子 不能保持完全的平行排列;另外,反密码子的第1个核 苷酸位于非双链结构的松弛环内,摇摆的自由度较大, 从而导致密码子的第3位核苷酸和反密码子的第1位核苷 酸之间形成非标准的碱基配对。(反密码子的这个位点 称为摇摆位点) 如果tRNA的摇摆位点是被修饰的碱基,就可能出现更 多的选择配对关系。
上次讲解内容
一、顺式作用元件与反式作用因子(重点) 二、真核生物RNA的转录过程 三、真核生物RNA转录后加工(重点) 1. 5’加帽; 2. 3’加尾; 3. 选择性剪接; 4. RNA编辑 四、RNA编辑
碱基的突变
C变为U
ApoB 基因有 29 个外显子
CAA
第 2153 个密码子编码 Glu 编辑
T-loop(TψC环)
• 这个环中始终含有胸 腺嘧啶-假尿嘧啶-胞嘧 啶的序列。 • 它与核糖体大亚基的 5S rRNA结合,稳定 蛋白质的结构
D-loop (DHU环)
直接与氨基酰tRNA合成酶
结合,使氨基酸连接到 tRNA的受体位点上。
tRNA与氨基酰tRNA合成酶的结合
氨基酸连接到受体位点上的过程:
UAA
3’UTR
AAA
Open reading frame(开放阅读框), ORF (3’非翻译区)
Stop codon(终止密码) UAG UGA UAA
开放阅读框(open reading frame, ORF): mRNA中从起始密码子(AUG)到终止密 码子(UAA、UAG或UGA)的核酸序列, 它可以编码一条完整的多肽链。
5蛋白质的翻译

proteins,r-proteins)组成,rRNA 组成总分子量的 60%~65%。核糖体的相对大小常常用 沉降系数单位来表示。大肠杆菌的核糖体称为 70S 核糖体,其中的小亚基称为 30S 核糖体, 大亚基称为 50S 核糖体(图 5-8) 。小亚基包含 21 种不同的蛋白质(被称为 S1 一 S21)和 16SrRNA。大亚基由 33 种蛋白质(被命名为 L1~L33)和 23S 及 5SrRNAs 组成(图 5-9)。真核 核糖体称为 80S 核糖体,其中 40S 小亚基包含 33 种蛋白质和 18SrRNA,而 60S 大亚基包含 50 种蛋白质(图 5-9)和 3 种 rRNA (28S, 5.8S 和 5S) 。 真核 5.8SrRNA 与细菌 23SrRNA 的 5SrRNAs 部分同源(表 5.1) 。古细菌核糖体类似于细菌核糖体,但有些包含与真核相同的特别亚基。
组织上,原核生物与真核生物有重要的差别(图 5.1、图 5.2 和图 5.3) 。原核生物的 mRNA 的第一个密码子 AUG 上游的一个重要特征就是 Shine-Dalgarno 序列,而真核生物 mRNA 除
第一个密码子 AUG 的上游是核糖体小亚基扫描 AUG 的信号序列(CCACC)外, 5’端非翻译区 上游为帽子结构, 3’端非翻译区内有多聚腺苷化的信号 AAUAAA 以及其下游的多聚 A 尾巴。 mRNA 是由 DNA 的模板链转录而来, 其序列与编码链相同与模板链互补。 mRNA 的 5’ →3 ’ 三联体密码子序列与蛋白质 N 端到 C 端的氨基酸序列线形相关。原核生物 mRNA 的转录和翻 译发生在时间与空间上具有相对的同一性,其 mRNA 通常不稳定,在合成后的几分钟内翻译 成蛋白质。 真核 mRNA 的合成与成熟都在核内, 成熟的 mRNA 被运往胞质, 作为模板翻译蛋白 质 , 其 稳 定 性 相 对 较 高 , 达 几 小 时 。
分子生物学-蛋白质的翻译课件

详细描述
核糖体通过识别mRNA上的起始密码子与mRNA结合,形成 翻译起始复合物。这个过程需要消耗能量,以确保核糖体正 确地定位在起始密码子上。
起始复合物的形成
总结词
起始复合物的形成是翻译过程的重要步骤,它涉及到多个蛋白质和RNA分子的相互作 用。
详细描述
起始复合物的形成涉及多个步骤。首先,核糖体与mRNA结合后,需要招募翻译起始 因子,如IF3和IF2。这些因子帮助核糖体正确地定位在起始密码子上,并确保翻译的准 确性。随后,氨酰-tRNA结合到核糖体的A位点上,准备开始多肽链的合成。至此,起
肽链的延长
01
02
03
肽键的形成
氨基酸在加入到肽链中后, 通过肽键的形成相互连接, 形成多肽链。
转肽酶的作用
转肽酶在肽键形成过程中 起催化作用,促进氨基酸 之间的连接。
核糖体的移动
随着肽链的延长,核糖体 沿着mRNA移动,确保下 一个密码子被正确识别和 翻译。
终止密码子的识别
终止密码子的种类
终止密码子有UAA、UAG和UGA三种,它们作为翻译终止的信号 被核糖体识别。
翻译的起始
02
起始密码子
总结词
起始密码子是mRNA上的一个特定 序列,用于标记蛋白质合成的起始位 置。
详细描述
起始密码子是mRNA上的三个连续的 核苷酸,通常为AUG。它不仅标记了 翻译开始的位点,还决定了从这里开 始合成多肽链的方向。
核糖体与mRNA的结合
总结词
核糖体是负责蛋白质合成的细胞器,它通过与mRNA的结合 开始翻译过程。
无意义校正是指当mRNA上的终止密码子提前出现时,核 糖体会提前终止多肽链的合成。这种机制有助于减少多肽 链的错误合成。
蛋白质翻译后修饰——末端修饰(氨基末端、羧基末端)——加密版

蛋白质翻译后修饰——末端修饰(氨基末端、羧基末端)(~~by luckyboy)(微生物班、精简打印、元旦巨献版)在核糖体上翻译的时候,当氨基酸添加到新生多肽之后,在体内氨基酸残基会发生各种各样的共价修饰。
I、氨基端的修饰初生蛋白的第一个氨基酸的命运:在细菌中:在细菌中生物合成蛋白质的第一步一般是甲酰甲硫氨酰-tRNAfmet和第二个氨酰tRNA通过肽键合成,因此初生蛋白质存在一个甲酰甲硫氨酰位点。
在真核生物中:虽然N末端甲硫氨酰位点从第一个甲硫氨酸获取在成熟蛋白质中很常见,N末端的α-甲基一般很快会被移除,接着在大多数情况下甲硫氨酸残基会被断裂下来。
这个作用是依靠甲硫氨酸氨基肽酶的作用,并且这个裂解过程由第二个残基控制。
(1)在酵母中(啤酒酵母):如果倒数第二个氨基酸残基有一个0.129nm或更小的回转半径的时候,甲硫氨酸会被完完全全的裂解掉(这些氨基酸有:Gly, Ala, Ser, Cys, Thr, Pro, Val)(3)在真菌或哺乳动物的线粒体中:起始甲硫氨酸的不被去除,但在植物的线粒体中还是会发生的。
在工程菌中:在大肠杆菌中过量表达的蛋白质通过质粒技术会导致一种甲硫氨酸残基保留的不正常现象。
一、乙酰化1.N端a-乙酰基修饰(a-acetyl)在蛋白质中是很普遍的在Ehrlich ascite 细胞中:大概有80%的可溶蛋白是N端a-乙酰基修饰的。
在高等真核生物中:有证据表明在这些细胞中氨基酸末端乙酰化是非常普遍的,几乎可以作为高等真核生物蛋白质的一个典型标志。
在低等真核生物中:N端a-乙酰基的比例比较低,但还是存在的2. N端a-乙酰基化修饰通常是翻译中同时发生的,一般发生在新生肽链大约40个残基长的时候3.N端残基乙酰化修饰的频率(概率)是不同的:一般Ala,Ser > Met,Gly, Asp > Asn,lle,Thr,Val >其他氨基酸残基(1)在高等真核生物中的蛋白质比细菌或真菌中的蛋白质更可能发生乙酰基修饰(2)在大肠杆菌中表达的真核细胞蛋白部分发生乙酰化。
5蛋白质翻译
AAAAAAA-OH
mRNA上存在遗传密码 mRNA分子上从5至3方向,由AUG开始,
每3个核苷酸为一组,决定肽链上某一个氨 基酸或蛋白质合成的起始、终止信号,称 为三联体密码(triplet codon)。
从mRNA 5端起始密码子AUG到3端终止
密码子之间的核苷酸序列,各个三联体密码 连续排列编码一条多肽链,称为开放阅读框 架(open
5' 7 m Gppp
reading frame, ORF)。
编码区 ORF AUG UAA 3' AAA……An
5'非翻译区
3'非翻译区
2、遗传密码的破译
• 基因密码的破译是六十年代分子生物学最辉煌的成就; • 1954年,物理学家George Gamow的数学推理(43=64 ); • 1961年,Brenner和Crick首先肯定了三个核苷酸的推理 ;
U
One tRNA anticodon may pair with as many as three codons.
3、遗传密码的特点
(1)连续性(commaless) :两个密码子之间无 任何核苷酸加以隔开和重叠,如插入/删除 碱基,可发生移码突变或框移
5′…. UACGGACAUCUG….3′
酪 插入 甘 组 蛋
5′….UACCGGACAUCUG….3′
酪 精 苏 半胱
缺失
5′…. UACGACAUCUG….3′
核苷酸结合技术
Nirenberg和Leder,1964
核苷酸结合技术