陕南奥陶纪圆尾虫类三叶虫的丰度及海平面变化
不屈行者——漫谈三叶虫的兴衰

46化 石2022年 第1期化石新知化石揭秘不屈行者——漫谈三叶虫的兴衰罗宜轩投名三叶虫(Tr i l o b i t e ),一般指节肢动物门(A r th ropoda )三叶形亚门(Trilobitomorph )三叶虫纲动物。
身体扁平,背侧被以坚固的甲壳,腹侧为柔软的腹膜和附肢。
背甲为两条背沟(dor salf u r row )纵向分为一个轴叶(axiallobe )和两个肋叶(pleurallobes ),前后两对附肢特化为触角和尾须。
身体自前而后又横分为头部(头甲)、胸部(胸甲)、尾部(尾甲)。
演替及其分析三叶虫作为古生代的标志性生物类群,地层年代中出现了两次发展的高潮期,第一次高潮期由三叶虫诞生始至奥陶纪末期生物集群灭绝事件止,第二次高潮期由志留纪末始至泥盆纪中期止。
第一次发展高潮期(约5.42亿前—约4.72亿年前)在寒武纪时期的海洋中,三叶虫拥有相对发达的身体构造,包括发达的复眼、坚硬的背甲,以及多对可以爬行的节肢步足(部分三叶虫发育有鳃状外肢,可能有仰泳的本领),因此在寒武纪的海洋中呈现出辐射适应,大约在4.9亿年前的寒武纪末期达到巅峰,达到了63个科,这个优势延续到了早奥陶世(统),并在约4.72亿年前结束。
奥陶纪早、中期继承了寒武纪时期环境特点,气候温暖,海侵广泛。
在4.9亿年前发生的浮游生物大爆发奠定了牙形动物繁盛的基础。
在约4.7亿年前,奥陶纪大辐射(Ordovician Radiation )的发生带来了角石类(Amenorrhea )等古生代演化动物群(Paleozoic Fauna )和板足鲎(亚纲)(Eurypterids )的兴盛,大量挤占三叶虫的生态位,导致三叶虫15个科灭绝,第一次发展高潮期结束。
同时,以三叶虫为代表的寒武纪演化动物群(Paleozoic Fauna )随后步入衰落。
自大氧化事件之后,地球大气的含氧量总体呈现上升趋势。
加之奥陶纪浮游生物革命(The Ordovician Plankton Revolution ),为动物的体型膨胀打下了基础。
三叶虫动物综述

三叶虫动物综述一、系统分类位置节肢动物门(Arthropoda)、三叶形亚门(Trilobitomorpha)、三叶虫(Trilobita)纲。
二、三叶虫的演化及地史分布三叶虫在寒武纪早期就已经出现。
其时已有4个目,属种丰富,全球分布。
并已显示出生物分区特性。
因此,很多学者认为三叶虫在寒武纪之前就已经存在。
从化石记录中可以分辨出其外部形态和身体构造在地质历史时期的演化趋势,分为4三、一般特征仅在古代的海洋中生活。
身体扁平,背侧为坚硬的甲壳,腹侧为柔软的腹膜、附肢。
自前而后分为头部、胸部、尾部。
又称头甲、胸甲、尾甲。
四、背夹(dorsal shield)构造1.头甲(cephalic shield):1.1.头鞍;1.2.颈环;1.3.颊部;1.4.眼叶;1.5.头盖;1.6.活动颊;1.7.面线;详见Figure 1,Figure 2,Figure 3。
Figure 1 眼Figure 2 头鞍Figure 3 头盖及活动颊2.胸甲(thoracic shield):2.1.轴叶;2.2.肋叶;2.3.半环;2.4.关节沟;2.5.间肋沟;2.6.肋沟;2.7.肋刺;2.8.关节面;详见Figure 4。
Figure 4胸甲3.尾甲(pygidial shield):半圆形。
若干体节融合而成。
第一个轴节上具半环和关节沟,以此和胸甲衔接。
根据头甲和尾甲的相对大小关系,将三叶虫分类为4种类型1)小尾型:尾甲极小。
2)异尾型:尾甲小于头甲。
3)等尾型:尾甲等于头甲。
4)大尾型:尾甲大于头甲。
球接子类(Agnostina)的头部无眼和面线,个体比较小,背夹基本构造和普通的三叶虫有所区别(Figure 5)。
Figure 5球接子五、三叶虫的分类由于这类动物已经灭绝,其内部构造及软体资料保存稀少,主要趋向于利用综合特点进行分类,主要考虑:○1形态特征;○2系统演化;○3地质地理分布。
六、三叶虫代表属1.球接子目Ptychagnostus:头鞍前叶亚三角形,基底叶被一对浅沟横穿。
三叶虫化石简介

三叶虫化石简介
三叶虫化石简介
三叶虫是繁盛于寒武纪(距今5.43亿年前)的无脊椎动物,其形态呈卵形或椭圆形,体长一般3—10cm,由前向后横分为三部分,即头甲、胸节和尾甲。
故名三叶虫。
三叶虫主要生活在浅海地带。
多数三叶虫以其扁平的身体贴伏在海底,向前作缓慢的爬行或作暂时的游移。
三叶虫盛极于寒武纪,从奥陶纪开始减少,直至二叠纪(距今2.5亿年前)绝灭,是受肉食鹦鹉螺类、鱼类的吞食,海水退缩,气候带变迁等因素的影响所致。
该景点位于景区北大门广场对面的黑山,此处的三叶虫化石俯拾即是,学名为中华莱得利基虫Redlichia chinensis。
产出的地层层位为下寒武统馒头组,是寒武纪早期的标准化石。
关于三叶虫有首诗说得好,头部半圆身体扁,胸节尾甲占半边,当年漫游浅海底,今日化石到人间。
三叶虫分类概说

三叶虫分类概说本文转自化石网,原作者铜钱豹三叶虫是生活在五亿至两亿年前的一种水生的小节肢动物,全国各地都有他们的踪影,很多猎人第一次猎石挖的就是三叶虫,他以他奇妙的结构与形态让许多猎人对他过目不忘,喜爱有加。
但是作为节肢动物,三叶虫的一大特点,就是分类繁杂,单属就有成百个,所以三叶虫的分类实在让许多人头疼不已。
那么如何对三叶虫进行大体的分类,今天我就介绍一种分类方法,这也是国际上现在比较通用的分类法:这种分发按结构将三叶虫分为几个目:AGNOSTIDA球接子目REDLICHIIDA莱德利基目CORYNEXOCHIDA耸棒头虫目LICHIDA裂肋虫目ASAPHIDA栉虫目PTYCHOPARIIDA 褶颊虫目ODONTOPLEURIDA齿肋虫目PROETIDA砑头虫目PHACOPIDA眼镜虫目HARPETIDA镰虫目这十个目看起来似乎非常难过繁杂,但是我们根据每个目的特点,就能把它们分开,下面我分别介绍一下AGNOSTIDA球接子目球接子目是三叶虫中最好辨认的一种了,它只要生活在寒武纪,尤其是寒武纪早期最为繁盛,这个目的三叶虫都有明显特点,1胸节少,大都只有两节。
最多三节2头尾等大。
如果被水流冲散很难分辨头尾3这个目大多数都有一个特点:没有眼睛,这是浮游生活的结果(如下图左边)而左图中的则是早期眼睛没有退化的一种球接子目代表种类:REDLICHIIDA莱德利基目莱德利基目是纯正中国制造,我国是这种三叶虫的发源地与大本营。
所以中国有世界上种类最多的莱虫种类,我们对莱虫也不太陌生,像云南的什么莱德利基,什么武定虫,都是这种。
这种三叶虫很好辨认1时间上它分布在寒武纪早起,中期就灭绝,所以中寒武纪以后没有莱虫。
2这个虫子的新月行眼睛是一大特色,又扁又长3尾巴很小,莱德利基虫是早期三叶虫,尾巴很小,几乎难以辨认,这也是他的一大特点莱德利基目代表种类:PTYCHOPARIIDA 褶颊虫目褶颊虫是所有三叶虫里面分类最为繁杂的,也是种类最多的,寒武纪是它的极盛时期,并发展为以后的各各目。
新疆塔里木盆地西缘奥陶纪上丘里塔格群三叶虫

新疆塔里木盆地西缘奥陶纪上丘里塔格群三叶虫
周志毅;周天荣;袁文伟
【期刊名称】《古生物学报》
【年(卷),期】1998(37)3
【摘要】首次记述了塔里木盆地西缘上丘里塔格群特马豆克晚期-阿仑尼克中期三叶虫3属4种,它们是深沟虫类Aksuaspisaksuensisgen.etsp.nov.,栉虫类Megalaspidesangustussp.nov.和M.sp.,以及1个属种不能确定的带针虫类。
三叶虫相和沉积相研究表明这一动物群生活于正常和风暴浪基面之间水动力能量较高的环境,代表一种低纬度带的浅水台地组合。
三叶虫具有强烈的地方性色彩。
三叶虫的组成和亲缘关系分析进一步证明塔里木陆块虽然古地理位置上靠近华南陆块,但应代表独立的板块。
【总页数】14页(P269-282)
【关键词】三叶虫;上丘里塔格群;奥陶纪;塔里木盆地西缘
【作者】周志毅;周天荣;袁文伟
【作者单位】中国科学院南京地质古生物研究所;西南石油地质局05项目工程处【正文语种】中文
【中图分类】Q915.2
【相关文献】
1.塔里木盆地北部上丘里塔格群致密灰岩微孔储层的发现及意义 [J], 叶德胜
2.塔里木盆地北部丘里塔格群(寒武系至奥陶系)白云岩的成因 [J], 叶德胜
3.巴楚地区上寒武-下奥陶统丘里塔格群碳酸盐岩储层特征 [J], 张新海;王晓东;阳国进;周新桂;毛晓长
4.塔里木盆地西缘上寒武统下丘里塔格群热液白云岩改造时限:来自古地磁的约束[J], 陈代钊; 张艳秋; 周锡强; 董少锋
5.新疆库鲁克塔格南部突尔沙克塔格群顶部(晚寒武世—早奥陶世)的三叶虫 [J], 林天瑞;张太荣;林焕令;周志毅
因版权原因,仅展示原文概要,查看原文内容请购买。
三叶虫的兴衰史
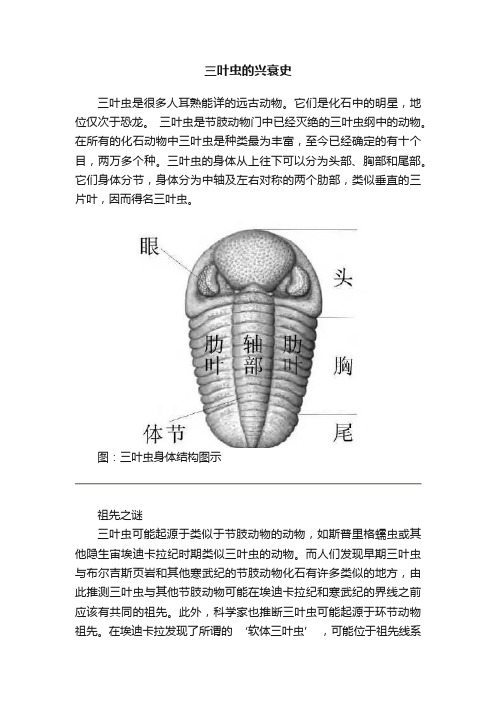
三叶虫的兴衰史三叶虫是很多人耳熟能详的远古动物。
它们是化石中的明星,地位仅次于恐龙。
三叶虫是节肢动物门中已经灭绝的三叶虫纲中的动物。
在所有的化石动物中三叶虫是种类最为丰富,至今已经确定的有十个目,两万多个种。
三叶虫的身体从上往下可以分为头部、胸部和尾部。
它们身体分节,身体分为中轴及左右对称的两个肋部,类似垂直的三片叶,因而得名三叶虫。
图:三叶虫身体结构图示祖先之谜三叶虫可能起源于类似于节肢动物的动物,如斯普里格蠕虫或其他隐生宙埃迪卡拉纪时期类似三叶虫的动物。
而人们发现早期三叶虫与布尔吉斯页岩和其他寒武纪的节肢动物化石有许多类似的地方,由此推测三叶虫与其他节肢动物可能在埃迪卡拉纪和寒武纪的界线之前应该有共同的祖先。
此外,科学家也推断三叶虫可能起源于环节动物祖先。
在埃迪卡拉发现了所谓的‘软体三叶虫’ ,可能位于祖先线系中的某个位置。
跌宕起伏早古生代(寒武纪、奥陶纪和志留纪)是三叶虫演化的鼎盛时期,这段时期三叶虫家族与地球历史发生若干次重大事件有密切关系。
晚古生代(泥盆纪、石炭纪和二叠纪)则是三叶虫的低谷。
我们接下来就以时间轴来揭示三叶虫家族一段高潮与低谷并存,繁荣与消亡兼有的辉煌历史。
寒武纪称雄寒武纪是三叶虫家族真正登上演化舞台的时期。
这个时候总体上三叶虫的头部比较大,尾部较小。
最值得一提的是三叶虫的眼睛。
大多数三叶虫眼睛具有连续角膜覆盖视觉区复眼;它们具有很多邻近的透镜紧密的堆积在一个视觉面上。
三叶虫眼睛的透镜体是由方解石构成的。
有矿物学知识的人可能知道方解石虽然透明但具有双折射率,不利于视觉系统。
但是三叶虫却良好地应用了这个系统。
图:中间型始莱德利基虫,产于云南澄江生物群(标本藏于南京古生物博物馆)最早粉墨登场的三叶虫种群是小油栉虫(Olenellina)家族,它们生活在海底。
后来出现了球接子目(Agnostida),耸棒头虫目(Cornexochida)和褶颊虫目(Ptychopariida)这些大家族。
奥陶纪

奥陶纪三字地理名词三字地质学术语中国地质摘要古生代第二个纪,约开始于5亿年前,结束于4.4亿年前。
在此期间形成的地层称奥陶系,位于寒武纪之上,志留纪之下。
奥陶纪是英国地质学家C.拉普沃思于1879年用Ordovices命名的,Ordovices 是威尔士地区的一古民族名“奥陶”一词系Ordovices的日文汉语音译。
奥陶纪包括原来属于A.塞奇威克命名的寒武纪和命名的志留系两地层的重复部分。
奥陶纪于1960年哥本哈根召开的第21届国际地质大会上正式通过。
奥陶纪-基本信息奥陶纪与头足类动物奥陶纪(Ordovician period,Ordovician)是古生代的第二个纪,开始于距今5亿年,延续了6500万年。
奥陶纪(Ordovician,符号O)是地质年代中的一个纪,开始于同位素年龄488.3±1.7百万年(Ma),结束于443.7±1.5Ma。
奥陶纪亦分早、中、晚三个世。
奥陶纪是地史上海侵最广泛的时期之一。
在板块内部的地台区,海水广布,表现为滨海浅海相碳酸盐岩的普遍发育,在板块边缘的活动地槽区,为较深水环境,形成厚度很大的浅海、深海碎屑沉积和火山喷发沉积。
奥陶纪末期曾发生过一次规模较大的冰期,其分布范围包括非洲,特别是北非、南美的阿根廷、玻利维亚以及欧洲的西班牙和法国南部等地。
奥陶纪-名字来源“奥陶”一词由英国地质学家拉普沃思(pworth)于1879年提出,代表露出于英国阿雷尼格(Arenig)山脉向东穿过北威尔士的岩层,位于寒武纪与志留纪岩层之间。
因这个地区是古奥陶部族(Ordovices)的居住地,故名。
奥陶纪-生物演化奥陶纪当时气候温和,浅海广布,世界许多地方(包括我国大部分地方)都被浅海海水掩盖。
海生生物空前发展。
化石以三叶虫、笔石(Graptolites)、腕足类、棘皮动物中的海林檎类(Cystoides)、软体动物中的鹦鹉螺类(Nautilites)最常见,珊瑚、苔藓虫、海百合、介形类和牙形石等也很多。
古生代的三叶虫

古生代的三叶虫古生代是地球历史上最古老的一个时期,距今约5.41亿年至2.51亿年。
在这个时期,地球上存在着众多神秘而奇特的生物,其中最著名的要数三叶虫。
三叶虫是一类古生代昆虫动物,其独特的外形和丰富的化石记录使其成为古生物学研究的热点之一。
本文将带你穿越时光,探索古生代的三叶虫世界。
一、古生代三叶虫的起源和进化古生代的三叶虫起源于晚寒武纪,经历了漫长的进化过程,最终繁荣于奥陶纪和志留纪时期。
作为节肢动物门三叶虫纲的代表,它们外形独特,通常被分为头胸部和尾部两个部分。
头胸部由多个肢节组成,每个肢节上都长有一对扁平的附肢,用于行走和觅食。
尾部则较为简单,主要用于平衡。
在古生代,三叶虫演化出了丰富的物种,不同物种具有不同的大小、体形和生活方式。
一些三叶虫物种生活在水中,生活习性类似现代的甲壳类动物,如螃蟹和虾类。
另一些三叶虫则生活在陆地上,具有较强的适应能力。
二、古生代三叶虫的生态角色古生代的三叶虫在当时的生态系统中起着重要的角色。
它们是海洋食物链中的重要环节,既是食物来源,同时也是捕食者。
三叶虫以浮游生物和底栖动物为食,通过剪取水中的浮游有机物维持自己的生存。
与此同时,一些大型的三叶虫也会主动捕食其他小型动物,成为海洋捕食者的一环。
此外,古生代的三叶虫也是地球生物多样性的重要组成部分。
它们属于脊足动物门,与今天的昆虫类动物有一定的关系。
通过对三叶虫化石的研究,科学家可以了解古生代生态系统的结构和演化过程,对地球生物多样性的起源和发展提供重要线索。
三、古生代三叶虫的化石记录古生代三叶虫的化石记录非常丰富,是古生物学研究的重要资源。
三叶虫的化石通常保存在沉积岩中,包括页岩、石灰岩和砂岩等。
这些岩石层一般形成于古代的海洋环境,因此三叶虫化石几乎遍布世界各地的古海洋沉积区。
通过对三叶虫化石的研究,科学家可以研究古生物的形态、生态和进化过程。
三叶虫的化石能够提供大量的信息,如体形、种类和年代等。
这些信息对于研究地球历史和生物演化具有重要意义。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
古生物学报,39(1):115-119(2000年1月)Acta P alaeontologica Sinica,39(1):115-119(Jan.,2000)收稿日期:1999 08 31*中国科学院南京地质古生物研究所现代古生物学与地层学开放研究实验室资助项目(No.913113)和国家自然科学基金资助项目(No.49772087)成果。
陕南奥陶纪圆尾虫类三叶虫的丰度及海平面变化*周志强(西安地质矿产研究所 西安710054)周志毅 袁文伟(中国科学院南京地质古生物研究所 南京210008)提要 上扬子区北缘米仓山地区3个宝塔组剖面的远岸水中浮游圆尾虫类三叶虫丰度变化研究表明,该区海域在兰维恩晚期至阿什极尔早期曾出现4次海平面跌落和3次海平面上升,与同期全球海平面变化有一定的可比性。
证明圆尾虫类丰度变化的研究可能是揭示奥陶纪全球海平面变化规律的一个有效手段。
关键词 圆尾虫类丰度变化 全球海平面变化 奥陶纪 陕西南部海洋生物群落的演替能反映海水相对深度的变化,运用浅水陆棚区的底栖生物群落或组合来识别海平面的变化,是近年国际上研究的热点。
笔者等近年来通过上扬子区北缘米仓山地区中 晚奥陶世宝塔组生物及生态地层学的初步研究,认为通过营远岸水中浮游(mesopelagie)生活的圆尾虫类(cy clopyg ids)三叶虫的丰度变化的研究也能有效指示海平面的升降,与陆棚区底栖动物群落研究有异曲同工的作用,应予重视。
圆尾虫类(cyclopygids)以其硕大的复眼和宽阔的中轴为特征,是繁衍于奥陶纪的一类特化三叶虫,营水中浮游生活,占据海洋的200 700m 深度带,分布在大陆斜坡的上部和有朝外海的自由通道的深水边缘盆地(Fortey,1985;Fortey and Ow ens,1987)。
周志毅等(1990)依据圆尾虫属(Cy clopyge )大量发现于新疆柯坪陆棚相其浪组之中,认为其生活水层的上限深度应浅于200m 。
经进一步研究,周志毅等(Zhou et al.,1994)指出圆尾虫类不同属虽然形态相似,但可能各自生活于不同深度的水层,即具有明显的深度分异现象,根据塔里木地区的实际资料,有些属如Microp ar ia (Micr op aria )和Cy clopyge 的生活水层深度上限可达100m 或更浅。
这一结论通过对滇西东部(印支地体)(Zhou et al.,1998)和上扬子区南缘奥陶纪三叶虫分布研究(Zhou et al.,1999)得到进一步支持。
从我们工作区的情况来看,圆尾虫类不同属的分布同样有深度分带现象,以Micr op aria 和Sagavia 为代表的组合,其分布范围可以从斜坡区一直延伸到浅外陆棚区的南郑元坝、南江桥亭一带;另一组合,包括Cy clop yge,Pricyclopyge,Microparia(Quadrata pyge ),Degamella,Asp idaeglina,Gastr op olus,Psilacella,Xenocyclop yge 则分布于斜坡区至深外陆棚区。
因此,应当说圆尾虫类总体上分布在深于100m 的水层中,其中绝大部分圆尾虫类的生活上限应浅于200m 。
既然圆尾虫类的分布严格受到深度控制,海水深度的变化必然会影响圆尾虫类在动物群中的相对丰度变化。
因此根据圆尾虫类丰度变化就可以识别海平面的升降。
工作区与上扬子区南缘(Zhou et al.,1999)一样,斜坡区的圆尾虫类在动物群中的相对丰度都在70%以上,而在浅外陆棚区则仅有零星出现,在位于这两个古地理单元的剖面上作圆尾虫类丰度变化研究显然不能灵敏反映海平面升降规律,因此我们在区内选择3个深陆棚相剖面即南郑梁山、胡家坝竹叶沟和大木沟剖面进行圆尾虫类相对丰度的统计。
这3个宝塔组剖面不仅从下到上均含圆尾虫类,而且它们的丰度变化显著,因而取得了较好的效果。
1 地质背景与时代扬子陆块北缘仅在陕西南部米仓山一带发育有较好的奥陶纪深外陆棚及斜坡相沉积。
笔者等对该区的宝塔组(包括原划的涧草沟组 见周志强等, 1999)进行了较详细的生物地层工作,依据圆尾虫类属种进行了三叶虫带的划分,同时也在杨铁汾的帮助下做了牙形石的分带工作。
该区在宝塔组沉积时期,在宁强 汉中一线以南海域属于浅外陆棚区;在勉县 大安一线以北,现今为碧口外来地体占据的地区,推测为深边缘盆地;二者之间则为深外陆棚 斜坡区(插图1)。
胡家坝镇附近的竹叶沟、大木沟剖面处于深外陆棚外缘,这里宝塔组的底界属于牙形石Pygodus serr a带之Eoplacognathus p rotor amosus 亚带的底,可与峡东地区宝塔组牯牛潭段上部至庙坡段下部同名亚带互比,时代为兰维恩晚期(王志浩等,1996)。
南郑梁山剖面则处于深外陆棚的内缘,宝塔组在此底界较高,属于牙形石H amarodus eu ropaeus带之底,与峡东地区宝塔组普溪河段(=原狭义宝塔组,见周志强等,1999)底界相当,时代为卡拉道克中期(杨铁汾,1996*)。
在大木沟剖面,宝塔组的顶部缺失;上述另外两个剖面,宝塔组的顶部则发育了原划为涧草沟组的N ankinolithus层,其时代为卡拉道克晚期 阿什极早期。
笔者在宝塔组划分了3个三叶虫间隔带(插图2),分带化石均是圆尾虫类属种。
Microparia(Quadratapyge)lep ida X enocyclopyge j askovitchi带以M icrop ar ia (Quadr atapyge)lep ida的首次出现为底界,以X enocy clopyge j askov itchi的首次出现为顶界,依据剖面中共生的牙形石或与剖面中牙形石带的上下关系分析,它大致与峡东宝塔组庙坡段牙形石Pygo dus anser inus带上部至Pr ioniodus alobatus带对比,属卡拉道克早期。
Xenocyclop yge j askovitchi Sagavia chuanx iensis带以X enocyclopyge jaskovitchi 的首次出现为底界,以Sagavia chuanx iensis的首次出现为顶界,它大致相当于峡东的宝塔组普溪河段,属卡拉道克中期。
Sagavia chuanx iensis Nankino lithus带则以Sagavia chuanx iensis的首次出现为底界,以Nankinolithus的首次出现为顶界,它大致相当于峡东地区宝塔组临湘段的主体,时代属卡拉道克晚期。
Nankinolithus组合带则可与峡东临湘段顶部的Nankinolithus带互比,为卡拉道克末期 阿什极早期。
2 圆尾虫类丰度变化曲线与海平面升降的关系在上述3个剖面中,化石样品采集的间距一般为0.5 2m,少量采集条件困难的地段间距放宽至3-5m。
绝大部分样品的采集量为10kg左右,采集的岩块又被剥离成厘米级的小块。
所含的三叶虫绝大部分呈头盖、活动颊、唇板、胸节、尾部分离状态保存。
为使统计的标本数量尽可能接近个体数插图1 陕南米仓山区中 晚奥陶世古地理和剖面位置图Sketch map show ing palaeogeography and location of M iddle Upper Ordovician sections in the M icangshan area of southern S haanxi*杨铁汾,1996:陕南宝塔组穿时性研究及奥陶系层序地层初探。
西安地质学院研究生学位论文。
116 古 生 物 学 报第39卷插图2 陕南米仓山区宝塔组圆尾虫类相对丰度变化曲线图Curves of relative abundances of cyclopygids through the Pagoda Formati on in the M icangshan area of southern Shaanxi量,同属一个种的标本只统计数值最大的分离部件,假如某一层位某种有100个头盖、15个活动颊、5个唇瓣、60个尾部,统计其数量为100个。
笔者统计了每一采集层位的圆尾虫类数量及三叶虫的总数量,然后依次算出每一层位圆尾虫类相对丰度的百分比值,并将剖面上各采集层位的丰度值连成丰度变化曲线(插图2)。
从插图2可见,3个宝塔组剖面圆尾虫类丰度变化曲线基本上是相似的。
在M icrop ar ia (Quadratapyge)lep ida X enocyclopyge j askovitchi 间隔带的下或中部、Xenocyclop yge jaskovitchi Sagav ia chuanx iensis间隔带的中部及Sagavia chuanx iensis Nankinolithus间隔带的下部分别出现了圆尾虫类丰度的3次高峰值(A1,A2,B1,B2,B3, C1,C2,C3)。
至于大木沟剖面在第二个峰值B2之上的次一级变化,则是由于该剖面化石采集密度较大所致。
相应的丰度低谷值则分别出现在牙形石Eo p lacognathus p rotoram osus亚带之上、三叶虫M i crop ar ia(Quadr atap yge)lep ida Xenocy clopyge jaskovitchi带顶部至X enocyclopyge j askovitchi Sagavia chuanx iensis带底部、Xenocy clopyge jaskovitchi Sagavia chuanx iensis带中上部及N anki nolithus带。
这种相似性显然不是偶然的,它说明在兰维恩末期 阿什极早期米仓山地区所属海域曾出现了海平面3次显著上升、4次显著跌落的过程。
为了便于和同时期的全球海平面升降变化曲线作比较,笔者等将3个剖面的圆尾虫类丰度变化曲线作了综合并放置在插图3的左侧。
该曲线在Sagav ia chuanx iensis Nankinolithus 带之下取自化石采集最充分的大木沟剖面,在Sagavia chuanx iensis Nankinolithus带至Nankino lithus带则主要依据竹叶沟的资料,以弥补大木沟剖面宝塔组顶部缺失的短处。
相应时期的海平面变化曲线则根据Ross和Ross(1992)关于北美、Fortey (1984)关于全球海平面变化的成果(插图3)。
两个海平面变化曲线在卡拉道克初期和末期出现的两次117第1期周志强等:陕南奥陶纪圆尾虫类三叶虫的丰度及海平面变化低谷分别对应于圆尾虫类丰度变化曲线在牙形石Eop lacognathus p rotor amosus 亚带和三叶虫M i cr op aria (Quadratapyge )lep ida X enocyclopyge j askovitchi 间隔带之间以及三叶虫Nankinolithus 组合带之内出现的2个低谷点。