第六章 植物胚与胚乳的发育
简述胚乳发育的模式及特点

简述胚乳发育的模式及特点胚乳发育是种子植物中胚乳细胞的发育和分化过程。
胚乳是种子中的重要组织,为胚胎提供营养和保护。
胚乳发育的模式和特点可以根据不同的植物物种进行分类。
1.油种类胚乳发育模式:油种类胚乳发育模式是最常见的一种类型,多见于豆科植物、蓖麻科植物等。
在种子成熟和胚乳发育的过程中,胚乳细胞会不断积累脂肪和蛋白质。
胚乳细胞中的储存蛋白质可以分为两类:一类是祖质蛋白质,主要用于胚胎发育所需;另一类是储存蛋白质,主要用于营养储备。
因此,油种类胚乳发育模式的胚乳细胞中富含大量的油脂和储存蛋白质。
2.淀粉种类胚乳发育模式:淀粉种类胚乳发育模式多见于禾本科植物、百合科植物等。
与油种类胚乳相比,淀粉种类胚乳中的胚乳细胞主要积累淀粉颗粒。
在发育过程中,胚乳细胞的蛋白质含量相对较低,而淀粉颗粒的数量和大小则显著增加。
淀粉种类胚乳发育模式的胚乳细胞中富含大量的淀粉,为胚胎提供能量和营养。
3.蛋白种类胚乳发育模式:蛋白种类胚乳发育模式多见于哺乳纲植物、银杏科植物等。
在这种模式下,胚乳细胞主要积累储存蛋白质,而油脂和淀粉的量较少。
蛋白种类胚乳发育模式的胚乳细胞中富含大量的蛋白质,为胚胎提供营养和保护。
胚乳发育的特点如下:1.营养积累:胚乳细胞在发育过程中不断积累营养物质,如脂肪、蛋白质和淀粉等。
这些物质为胚胎提供了所需的能量和营养。
2.组织分化:在胚乳发育的过程中,胚乳细胞会分化为不同的类型,形成不同的组织。
油种类胚乳常由内外子皮、胚乳皮和胚乳核等组成,而淀粉种类胚乳则由胚乳皮和胚乳核组成。
3.储存功能:胚乳细胞在发育过程中不仅能够积累大量的营养物质,还能将其储存和保护起来,以供胚胎发育和新植株的生长所需。
4.营养转移:在种子萌发过程中,胚乳细胞会释放出储存的营养物质,供胚胎和新植株生长所需。
这个过程被称为乳沟。
总的来说,胚乳发育在种子植物的生殖过程中起着重要的作用。
它不仅为胚胎提供所需的能量和营养,还起到了保护和储存的作用,对于新植株的生长和繁殖至关重要。
第六章 植物胚与胚乳的发育
由运输载体在质膜上的数 量和分布控制
Chapter 8 Embryogenesis and seed development
8.1 Embryogenesis
Chapter 8 Embryogenesis and seed development
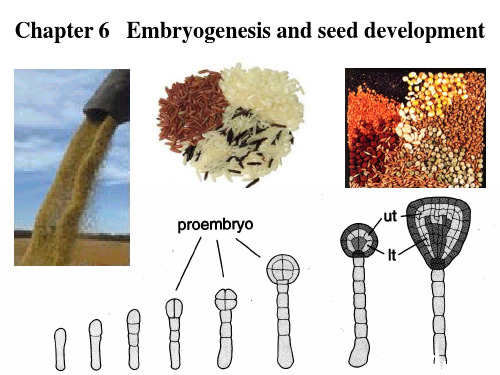
在球形胚的晚期,胚根原细胞发生不 对称分裂,形成一个小的顶细胞和一 个较大的基细胞; 顶细胞继续垂周分裂形成两个小细胞, 组成QC; 基细胞继续分裂形成小柱干细胞 子叶在球形期晚期开始形成
What activities are happened in embryogenesis? 胚胎发生过程中的事件
13kinesin微管1415ef为普通荧光显微latb生长素运输抑制剂褐藻黑角菜海藻胚发育过程中的肌动蛋白分布16顶轴的形成合子不对称分裂后分裂成基轮细胞分裂后16细胞期顶细胞形成原胚基细胞形胚根原细胞参与ram的形成中是qc和小柱细胞的原始细胞细胞期16细胞期17
Chapter 6 Embryogenesis and seed development
Origin 2.2 Origin of cellof polarity cell polarity
Different distribution of mRNAs, proteins and organelles between two poles 在极性细胞两极分布 着mRNAs,蛋白质 和不同的细胞器
GCm
5‘ direct repeats The 5' direct repeats of the FWA gene
Regulator Exon intron
Enhancer
Promoter
DNA methylation
胚乳、胚和种子的发育

小麦的胚乳在授粉后48—50h,游离核为100 个左右时开始形成细胞壁。 锦葵属、苹果属、芸苔属和柑桔属在形成数 百个至数千个游离核时才形成细胞。
多数植物的胚乳在发育后期才形成细胞
随着游离核的增加和液泡的扩大, 核常常被挤到周缘,并多集中在 珠孔端和合点端,胚囊中央被一 个大液泡占据。 游离核胚乳形成细胞壁的顺序, 一般是从珠孔端向合点端,从胚 囊周边向里面自由地推进。
三、沼生目型胚乳
初生胚乳核分裂— 珠孔室:大,游离核分裂—胚 乳细胞 合点室:小,分裂少,保持游 离核状态
第二节
胚的发育
受精的卵细胞称为合子(zygote),胚的 发育始于合子,经过原胚和胚的分化发育 阶段,最后达到成熟。
一、合子
要经过休眠期才进行分裂,故合子的分裂一 般迟于初生胚乳核。 休眠期的变化:极性加强 ,体积缩小 、增 大 、显著地伸长 ,细胞质中各 种细胞器数量增加并重新分布 为代谢活跃的细胞。
二、原胚
从合子的第一次分裂形成的两细 胞开始,至器官分化前的胚胎发育阶 段,称为原胚时期。
合子——顶细胞:多次分裂形成胚体 基细胞:形成胚柄
荠菜胚的发育是双子叶植植物胚的发育
2 单子叶植物胚的发育 裂叶茨藻(Najas lacerata)
传递细胞型的特征几种植物早期的胚示胚柄吸器种子是由胚珠经过双受精作用后发育形成的是胚这一新孢子体发育的场所
第十章
胚乳、胚和种子的发育
第一节 胚乳
精核与二极核融合后,一般不经过休眠,很快 便开始初生胚乳核的分裂,比合子的分裂早得多
一、核型胚乳 核以游离的状态分布在中央细胞的细 胞质中。胚乳维持游离的核状态的时期 长短因植物而异。
种子的发育、果实的形成及果皮的结构 (植物学)(共86张PPT)

〔1〕肉果〔fleshy fruit〕
① 浆果(berry): 由复雌蕊的上位子房或下位子房发育而成。 外果皮薄,中、内果皮多汁,皆肉质化。 如葡萄、番茄、柿、香蕉等的果实。
② 核果(fleshy fruit)
由单雌蕊或复雌蕊的上位子房或下位子房发育而成,具有坚硬 果核。
中果皮疏松髓质,内果皮膜质,分为假设干室,室内充满含汁的长形丝状 细胞,由子房内壁的毛茸发育而成。 ② 核果:通常由单雌蕊发育而成,内含一枚种子,三层果皮性质不同。 ③ 梨果:子房下位,果实由花筒和心皮局部愈合形成。
〔2〕干果:果实成熟后,果皮
① 裂果:果皮开裂
枯燥
A、荚果:单心皮发育而成,两个缝线开裂
沿心皮〔或子房室〕间的隔膜开裂,但子房室 的隔膜仍与中轴连接, 如牵牛等的果实;
孔裂( porous dehiscence):
果实成熟,心皮上方裂成一个小孔,种子由小孔中因风吹摇动而 散出,如虞美人、金鱼草的果实;
盖裂(circumscissile dehiscence):
果实成熟后,沿果实的中部或中上部作横裂,成一盖状 脱落,
第一节 种子的发育
种子的形成
种子的形成:包括胚,胚乳和种皮的 形成。
它们分别由受精卵,受精的极核和珠被 发育而来。
胚珠 → 种子
胚囊
受精极核 → 胚乳
胚珠 珠心〔吸收〕受精卵 → 胚
珠被
→ 种皮
种子
一、胚的发育
由受精卵发育为胚的过程中要经过 两个阶段:休眠阶段和原胚阶段
休 眠:卵受精后,产生一层纤维素的壁, 便进入休眠状态。休眠期的长短,一般 几小时,也有的长达几个月。
〔2〕非纯由子房发育而成——假果〔南瓜, 苹果〕
第六章:胚乳培养

胚乳培养中存在的问题
从染色体数目的分析表明,胚乳愈伤组 织及胚乳植株染色体数目变化很大。如 在苹果“金冠”和“红玉”的胚乳植株 中,其根尖染色体数目分别为32~48和 27~58左右,其中多于35条非整倍体细 胞十分普遍。而直正三倍体的细胞却很 少。在大麦的胚乳培养中根尖染色体数 目可以从1到4倍,另外还有许多具6、8、 11、12、13条染色体的非整倍体细胞。
胚乳培养中存在的问题
此外,还可以从理论上探讨引起倍性混 乱的原因及其控制的可能性,研究胚乳 植株形态发生规律及其遗传性状变化问 题以及胚和胚乳的关系等等。
胚乳培养中应注意的问题
1. 确定适宜的发育时期
胚乳的发育,大致可以分为早期、旺盛生长期 和成熟期三个阶段。处于发育早期的胚乳,如 核型胚乳的游核时期或刚刚形成细胞时期的胚 乳,难于培养产生愈伤组织或形成器官。生长 旺盛期的胚乳,大多是取材的最合适时期,以 前培养成功的植物,大都是取这个时期的胚乳, 如大麦、水稻、苹果、猕猴桃等。少数是取成 熟时期的胚乳也能获得成功,如蓖麻、巴豆、 麻风树、罗氏核实木等。
胚乳培养中应注意的问题
植物种类不同,胚乳生长旺盛阶段的日期也不 会一样,此时的胚乳具有:胚已分化完成,胚 乳已充分生长,几乎达到成熟时大小;外观为 乳白色半透明的固体,且富有弹性等特点。在 开始对任何一种植物进行胚乳培养时,除了要 查阅有关文献外,还应事先按期采集和固定开 花的材料作为系统观察,以确认该植物胚乳的 发生类型和发育时期,为下一步深入工作打好 基础。
胚乳的发育过程

二、胚乳的发育过程
禾本科植物和拟南芥胚乳发育经过两个主要阶段:多核体 胚乳阶段和胚乳细胞化阶段。
多核体胚乳阶段中三倍体胚乳核经过多次分裂,核之间不 形成成膜体,游离的核先在中心细胞珠孔端均匀分布,然 后在中心液泡外层细胞质中均匀分布,直到整个细胞质中 平均分布,成为完全的胚乳多核体。
Betl1(Basal endosperm transfer cell layer1)在胚乳基部细胞 表达,与细胞壁紧密结合。MEG1是富含半胱氨酸的糖蛋白, 胚乳发生早期在传递细胞中只有母方基因表达,父方基因被 甲基化失活,晚期父母双方的基因都表达,说明母方印迹在 早期的作用 。
BAP2胚乳基部细胞之间,主要积累在花梗厚壁的细胞间。 此外,相关区域表达的蛋白还有玉米BETL2、BETL3、 BETL4、BAP1 (BASAL LAYER-TYPE ANTIFUNGAL PROTEIN1)、BAP3。许多蛋白与抗微生物蛋白相似,说 明具有防御功能 。
种子大小的决定
二倍体母体的珠被、单倍体胚囊、三倍体胚乳以及胚之 间在发育过程中发生相互作用,共同决定种子的大小。
一、母体珠被影响胚乳的发育和种子大小
拟南芥珠被细胞繁殖的起始、最终细胞数目与细胞延长平衡, 不由种子大小决定。珠被细胞延长在胚乳大小与珠被之间的 协同中起关键作用,这种协同影响以后的胚乳细胞化和胚细 胞繁殖,最后决定种子大小。
体外玉米胚乳器官培养研究表明,胚乳细胞的与母体相邻的 表面位置是决定糊粉层分化的充分因素。
培养的胚乳组织外层糊粉层细胞分裂后形成的内层子细胞逐 渐失去传递细胞特性,dek1和sal1突变体胚乳组织在体外培 养同样表现出没有糊粉层和多层糊粉层的表型。
DEK1和CR4在其他拟南芥其他器官表皮中也起着决定表皮 特性的作用,说明表面位置是决定糊粉层形成的重要因素 。
第六章植物胚培养ppt课件
植物组织培育
第六章 植物胚培育
大 家 好 !
刘忠德 13082772026
第六章 植物胚培育
植物胚培育 是指在无菌条件下,对植物的胚 及胚器官如子房、胚珠和胚乳进展离体培育 的技术。
1904年Hanning进展萝卜 和辣根菜成熟胚培育
胚培育的类型和意义
本章内容
胚培育的方法
常见植物胚的培育
本章教学目的与要求
(1)了解植物胚培育的类型 (2)掌握胚培育的方法和操作步骤 (3)了解常见植物胚培育的技术
〔3〕多数植物成熟胚的生长以12h/天光照为宜。
牡丹成熟胚的离体培育与快速繁衍
牡丹成熟胚的离体培育
以‘凤丹白’成熟胚为外植体 结果阐明: 〔1〕突破成熟胚休眠最适宜的预处置是4℃沙藏。 〔2〕成苗培育时的最正确的培育基是MS+6-
BA0.2mg/L+IBA1.0mg/L。 〔3〕最适宜的生根培育基是MS+IAA1.0mg/L 。
1、胚珠培育 定义:是指将胚珠从母体上
分别出来,在无菌的人工环 境条件下培育,使其生长发 育构成幼苗的过程。
倒生胚珠
2、胚珠培育类型:
〔1〕、受精胚珠的培育 〔2〕、未受精胚珠的培育
示胚珠的位置
3、胚珠培育的根本过程:
〔1〕培育基:White,Nitsch,MS培育基
〔2〕胚珠的发育: A 受精胚珠:一是构成种子;二
柑橘:珠心组织 发生多个不定胚。
第六章植物胚和胚乳的发育
第六章植物胚和胚乳的发育
二、胚乳的吸器结构
• 豆科和葫芦科植物,存在胚乳吸器是十分普遍的。 在已研究的豆科20种植物中,均发现有合点吸器。
• 在胚发育至球形之前,珠孔端的胚乳就向心地形 成细胞,细胞的胚乳只局限在胚囊的上半部或三 分之二的部分,一般称之为胚乳本体,下部则维 持游离核的状态,成为多核的吸器。
第六章植物胚和胚乳的发育
第六章植物胚和胚乳的发育
2 糊粉的和淀粉的胚乳的分化
• 胚乳的最外层的特殊形态和生理性质,在禾本科 植物中早已引起重视和进行过许多的研究。小麦、 大麦和玉米等禾本科植物中,胚乳组织的表面层 细胞被称为糊粉层。它区别于内部胚乳细胞,最 明显的是含糊粉粒而不是含淀粉粒。这两种性质 不同的胚乳组织的形成,是从细胞化过程中第一 次平周分裂开始。此次分裂产生的周缘子细胞为 糊粉胚乳的原始,而内部的小窝及其继续的平周 分裂产生淀粉的胚乳细胞。这两种细胞其后的分 裂面是不同的,糊粉细胞进行平周分裂,分裂面 和成膜体由早前期微管带预定,而淀粉细胞则缺 少早前期微管带,分裂面是无规律的
• 受精后在胚乳和胚生长的同时,胚囊扩大,珠心 自溶。一般认为珠心自溶是被胚乳吸收作为自己 的发育。
• 有些植物在受精前中央细胞的珠孔端或合点端的 壁具传递细胞型壁内突的特点,受精以后壁内突 继续存在。此时,壁的内突侵入胚乳,由于质膜 表面积扩大,有利于营养物质从母体组织运输到 胚乳。
• 在种子发育中,吸器从母体组织获取营养无疑起 着重要的作用。
• 裸子植物的种子中也具有胚乳,但其从来源上讲 是雌配子体的延续,是单倍体。被子植物的胚乳 是由一个精子和两个极核融合产生的,一般是三 倍体,也有一些特殊情况,如百合属和贝母属是 五倍的,另外有少数的科的植物在胚囊发育的过 程中几乎不形成胚乳。
植物坯胎培养及离体授粉
离体授粉的方法: 试验材料的选择:以子房较大胚珠较多的植物较易成功。
确定开花、花药开裂及授粉时间
壹
贰
去雄后将花蕾套袋隔离
叁
制备无菌子房或胚珠
肆
制备无菌花粉
伍
胚珠或子房的试管内授粉
离体授粉的程序:
01
外植体
03
培养条件
02
培养基
影响离体授粉受精的因素:
为什么幼胚培养比成熟胚培养要求的培养基和培养条件更为严格?
未授粉子房培养形成小的无籽果实。
子房培养:
未授精子房培养为试管受精提供技术基础;
未受精子房培养能获得获得单倍体植株,用于单倍体育种。
获得杂种植株;
子房培养的意义:
培养基:基本培养基(MS、N6、B5等)、植物激素(配合使用)、蔗糖浓度(3-10%)
培养方式:直插优于平放。单倍体胚有多种,可以是卵细胞、助细胞或反足细胞等。
碳水化合物:
胚发育早期要求较高的的渗透压,而随着胚的发育逐渐降低。
成熟胚一般不需要外源激素即可萌发。但激素能促进休眠胚的萌发。
幼胚需要外源激素,应保持外源激素与内源激素的平衡。过低则不能促进生长,过高则易导致幼胚脱分化形成愈伤。
02
01
植物激素的作用:
天然提取物及某些蛋白制品;
银杏幼胚培养中添加银杏胚乳有促进作用(1934);曼佗罗幼胚培养中发现添加椰子乳能很好的促进幼胚的发育(200-500um的幼胚) (1941)。
未授粉胚珠:开花前1-6天,
授粉胚珠:授粉后1-120天不等,较晚时期有利于胚的发育。
灭菌:先对子房或外表组织消毒灭菌后,后取出胚珠培养。
01
03
02
04
植物胚与胚乳
1.胚与胚乳的形成1.1 被子植物的卵细胞、极核、精子的形成了解被子植物的卵细胞、极核、精子的形成过程是很好解决胚和胚乳基因型问题的基础。
孢子减数分裂(居间减数分裂)是其形成卵细胞、极核、精子的方式,具体过程为:花药中产生小孢子母细胞(花粉母细胞)四分孢子4个小孢子(花粉粒)4个成熟的花粉粒(各有1个营养核和1个生殖核)8个精子(每1个花粉粒萌发产生2个具有相同基因型的精子)。
胚珠中产生大孢子母细胞(胚囊母细胞)4个大孢子(其3个退化,1个发育)8核胚囊(1个卵细胞、2个极核、2个助细胞和3个反足细胞的基因型相同)。
1.2 被子植物的双受精当花粉落在柱头上,受到柱头上黏液的刺激,开始萌发,生出花粉管,花粉管穿过花柱,进入子房一直达到胚珠,同时放出两个精子,一个精子与卵细胞融合,一个精子与两个极核融合,这样两个精子分别与卵细胞和极核相融合的现象,叫做双受精作用。
双受精所形成的胚核和胚乳核经过发育分别形成胚和胚乳。
1.3 胚与胚乳基因型的组成胚基因型为卵细胞与精子基因相加组成,胚乳基因型为两个极核与精子基因相加组成。
2.胚与胚乳基因型推理、判断的一般思路与方法先根据亲本基因型推断其精子、卵细胞(极核与卵细胞相同)的基因型,然后根据双受精的原理写出种子的胚及与之对应胚乳的基因型,再验证之。
如:例1、如果某种植物的基因型为AaBb,这两对基因独立遗传,则其胚及胚乳可能的基因型是:()A、AaBb、AaaBBbB、AaBb、AaabbbC、AABb、AaaBBbD、AABb、AAabbb解答过程:①根据亲本推精子、卵细胞及极核基因型为:精子为AB、Ab、aB、ab,卵细胞(极核)基因型为AB、Ab、aB、ab。
②根据精子、卵细胞及极核基因可写出胚及其与之对应的胚乳基因型(表1)表1 由精子、卵细胞及极核基因型推导胚及胚乳基因型卵细胞(极精子胚胚乳结果答案核)ABAB AABB AAABBBAb AABb AAABbb aB AaBB AaaBBB ab AaBb AaaBbbABAb AABb AAABBbAb AAbb AAAbbbaB AaBb AaaBBb 可能 A ab Aabb AaabbbABaB AaBB AAaBBBAb AaBb AAaBbb aB AaBB AaaBBB ab AaBb AaaBbbABab AaBb AAaBBbAb Aabb AAabbbaB AaBb AaaBBbab aabb aaabbb例2、如果某种植物的基因型为AABb和Aabb,控制不同性状的两对基因分别独立遗传,则其种子的胚和胚乳基因型可能是:()A、AABb、AAAbbbB、AABB、AAABBbC、AaBb、AaaBbbD、aabb、AaaBbb解答过程:①根据亲本求精、卵细胞(极核)基因型为:精细胞为AB、Ab或Ab、ab,卵细胞(极核)基因型为Ab、ab或AB、Ab。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
in Arabidopsis
RT-PCR analysis of FWA expression in various organs 分析FWA在不同器官中的表达
FWA::FWA-GFP fusion protein analysis
FWA基因在受精后的种子和胚乳中表达
Endosperm analysis with Confocal
•Polarity establishment 极性的建立(合子)
•Cell fate 细胞命运决定(合子
经过胚胎发育过程分化出了性质不同的 、处于特定位置的细胞群,并形成了特 定的器官原基和组织)
Ut,外层 It, 内层
顶细胞分裂到8细胞期,外层细胞特化为表皮原细胞,内 层细胞特化为形成层和维管束原细胞
Regulator
Exon
Promoter Enhancer
intron
DNA methylation mCG
GCm
XY
XX
5‘ direct repeats The 5' direct repeats of the FWA gene
Regulator
Exon
Promoter Enhancer
intron
1. 受精 2. 孤雌生殖 3. 花粉粒 4. 愈伤组织
Egg
Ovule and embryo sac
胚胎发生: 从受精卵(合 子)发育成胚
的过程
双子叶植物拟南芥的胚胎发育过程
伸长期 合子经过 四 合子 第一次分 分 裂,产生 体 1个顶细 胚 胞(绿色)
和一个基 细胞(粉红
色)
16
早
心
成
细
期
形
熟
胞
心
胚
胚
球
形
形
胚
胚
1:顶细胞;2:基细胞;3:胚根原细胞,形成未来的 ROC和根冠中央区;RM:根尖生长点;SM:茎尖生长点
What activities are happened in embryogenesis? 胚胎发生过程中的事件
What activities are happened in embryogenesis? 胚胎发生过程中的事件
玉米和拟南芥的胚乳中均已发现印迹基因
玉米:R基因
拟南芥:MEA、FWA
m: maternal(母本); p: paternal(父本)
营养生长 阶段
配子体发生
花粉(雄配子体)
拟南
芥中
MEA
基因
组印
受精
迹发
生模
式图
组蛋白H3K27甲基化
Why endosperm-specific?
Studies on the gene
合子对称分裂
The polarity along apicalbasal axis
is disordered.
Pattern mutations
胚胎发育的模式建成包括不同方向上极性建立的机理
生长素的转运在胚胎发生中起着重要的作用 32细胞球形胚阶段
胚轴形成与生长素转运
生长素流的强度与方向只 由运输载体在质膜上的数
Egg
Polarity in single cells
Or2i.g2inOorfigcienllopfoclealrliptyolarity
Different
distribution of
mRNAs, proteins
Egg
and organelles
between two poles
在极性细胞两极分布 着mRNAs,蛋白质
WOX (WUSCHEL related homeobox)
(1)基-顶轴极性中 WOX2/WOX8,9 调控途径
WOX2和WOX8在合子中共表达; 在发生不对称分裂后,WOX2仅 在顶细胞中表达;而WOX8/9在
顶端胚柄细胞中表达;同时 WOX8的mRNA可以在胚柄下端
细胞中检测到
(2)外层细胞和内层细胞调控基因
在球形胚的晚期,胚根原细胞发生不 对称分裂,形成一个小的顶细胞和一 个较大的基细胞; 顶细胞继续垂周分裂形成两个小细胞, 组成QC; 基细胞继续分裂形成小柱干细胞
子叶在球形期晚期开始形成
What activities are happened in embryogenesis? 胚胎发生过程中的事件
development including FIE,FIS1-2
(2) 胚乳发育中的基因组印迹:
基因组印迹是一种非孟德尔遗传现象。它是指在配 子或合子发生期间来自亲本的等位基因或染色体产 生专一性的加工修饰导致后代体细胞中两个亲本来 源的等位基因有不同的表达活性,又称遗传印迹或 亲代印迹或配子印迹。
•Polarity establishment 极性的建立(合子)
•Cell fate 细胞命运决定(合子
经过胚胎发育过程分化出了性质不同的 、处于特定位置的细胞群,并形成了特 定的器官原基和组织)
•PCD 细胞程序性死亡(基细胞
分裂产生的子细胞,除胚根原细胞之外 ,其它所有细胞均不参与胚的形成,而 是共同构成胚柄,将胚胎与珠孔端的母 体组织连在一起,并在心形胚后期开始 进入程序化死亡而退化)
What activities are happened in embryogenesis? 胚胎发生过程中的事件
•Polarity establishment 极性的建立(合子)
•Cell fate 细胞命运决定(合子
经过胚胎发育过程分化出了性质不同的 、处于特定位置的细胞群,并形成了特 定的器官原基和组织)
Chapter 6 Embryogenesis and seed development
Chapter 8 Embryogenesis and seed development
8.1 Embryogenesis 胚胎发生 8.2 Endosperm development
胚乳发育
植物胚的不同来源途径
胚乳的发育
胚乳发生:中央细胞 极核与精细胞融合。
(1)胚乳发育模式:无需双受精的胚乳发育
没有受精而自主进行 种子发育的fis突变体
没有受精的FIS植株 角果
受精的FIS植株 角果
?
Why fertilization is needed for endosperm development?
为什么双受精是胚乳发育所必须的呢? Are there endosperm-specific genes? 胚乳发育特异
DNA methylation
Plus Biotin in medium
(包含生物素的培养基)
Lack of biotin
bio
Plus Biotin in medium
raspberry:
Wild type 球形胚无法转变
Other stage-specific genes 为心形胚
Twin mutant 双胚突变体
Disorder in PCD?
Embryo –lethal mutants in maize (玉米)
emb
?
突 变
Mutant bio in
体
Arabidopsis
胚的发育受到影响
Embryo –lethal mutants in maize
A mutant in Arabidopsis
Lack of biotin
(生物素缺乏)
1) Embryo-lethal mutations 2) Pattern mutations
模式突变体
Pattern mutations
Pattern mutations
The polarity along apicalbasal axis
is disordered. 丧失了极性
And the zygote divides symmetrically.
temporally expressed in endosperm?
DNA 甲基化 分析
受精前在中央 细胞中表达
受精后6小时 受精后12小时
中央细胞
受精后24小时
受精后48小时 GFP无法检测到
DNA methylation mCG
胚乳中DNA甲基化程度偏低,发生去甲基化
GCm
5‘ direct repeats The 5' direct repeats of the FWA gene
•PCD 细胞程序性死亡(基细胞
分裂产生的子细胞,除胚根原细胞之外 ,其它所有细胞均不参与胚的形成,而 是共同构成胚柄,将胚胎与珠孔端的母 体组织连在一起,并在心形胚后期开始 进入程序化死亡而退化)
•Regulation on gene expression基因表达调控
发育特异基因 (Developmental specific genes)
共聚焦显微境观察FWA基因表达模式
Why FWA only
受精前在中央 细胞中表达
受精后6小时 受精后12小时
temporally
expressed in
endosperm?
中央细胞
受精后24小时
受精后48小时 GFP无法检测到
Endosperm analysis with Confocal
Why FWA only
的基因?
Why endosperm-specific?
fie mutant: endosperm developed without fertilization (Ohad et al.,1999)
At least 3 gFenIEesecnocnotdroesl eanpdoolsypceormmb-inpdreopteeinndent