多细胞动物的起源学说
第4章_多细胞动物的起源0
两侧对称动物:其他所有门类
后生动物在胚胎发育过程中有胚层的分化, 多孔动物门只有内胚层和外胚层的初步分化, 腔肠动物门在内外胚层间又有中胶层。 自扁形动物门以后的门类都是三胚层动物。
根据体腔的有无和结构可将后生动物分为 无体腔动物:包括多孔动物门、腔肠动物门和 扁形动物门; 假体腔动物:包括线形动物门、腹毛动物门等; 体腔动物:包括环节动物以后的所有动物门类。
4.1 从单细胞动物到多细胞动物
单细胞动 物(原生 动物)
中生动物
多细胞动 物(后生 动物)
侧生动物 真后生动物
4.1 从单细胞动物到多细胞动物
进化顺序
原生动物→中生动物→后生动物
中生动物(双胚虫、异胚虫)
小型内寄生动物 结构简单 约50种 细胞数目恒定 其分类位置尚无定论
后生动物是除原生动物外所有其他动物的 总称(后生动物亚界)。动物界除原生 动物门以外的所有多细胞动物门类的总 称。
真后生动物
动物界除原生动物门以外的所有多细胞 动物门类的总称。 其特征是体躯由大量形态有分化、机能有分 工的细胞构成; 与群体原生动物的兼有营养和生殖功能的细 胞不同,其生殖细胞和营养细胞有明显的分 化。
真后生动物
依体制形态的对称情况,后生动物: 不对称动物:多孔动物门
辐射对称动物:腔肠动物门、栉水母动物门、 棘皮动物门
4.2 多细胞动物起源于单细胞动物的证据
一、古生物学 地层中动物化石
如:最古老的地层化石最简单,晚近的地层动物化 石种类多且复杂。
二、形态学 单细胞→多细胞,简单→复杂,低等→高等
如:原生动物的团藻等群体形态与多细胞动物相 似,可能为中间类型,以群体的形式过渡。
三、胚胎学 多细胞动物从受精卵开始,经卵裂、囊胚、原肠
多细胞动物的起源学说

多细胞动物的起源学说(一)群体学说大多数学者认为,多细胞动物起源于群体鞭毛虫类似的祖先。
对此也有两种假说:1、原肠虫学说赫克尔提出和团藻相似的群体单细胞动物一端内陷,形成了有原肠和两胚层的原始多细胞动物。
把此祖先称为原肠虫。
2、吞噬虫学说梅契尼可夫提出,具有单层细胞的单细胞群体内,一部分细胞摄取食物后进入群体之内,形成了两胚层的实心的原始多细胞动物。
把此祖先称为吞噬虫。
因现存的较低等的动物,多由细胞移入而形成两胚层。
内陷法到后来才有。
因此,吞噬虫学说可能更接近于事实。
(二)合胞体学说认为多细胞动物起源于多核纤毛虫的原始类群,后生动物的祖先是具合胞体结构的多核细胞。
生物发生率(biogenetic law)也叫重演律(recapitulation law),是德国人赫克尔(E.H.Haeckel)用生物进化论的观点总结了当时胚胎学方面的工作提出来的。
当时在胚胎发育方面已揭示了一些规律,如在动物胚胎发育过程中,各纲脊椎动物的胚胎都是由受精卵开始发育的,在发育初期极为相似,以后才逐渐变得越来越不相同。
达尔文曾作过一些论证,认为胚胎发育的相似性,说明它们彼此有亲缘关系,起源于共同的祖先,个体发育的渐进性是系统发展中渐进性的表现。
达尔文还指出了胚胎结构重演其过去祖先的结构,“它重演了它们祖先发育中的一个形象”。
海克尔明确地论述了生物重演律。
1866年他在《有机体普通形态学》书中说:“生物发展史可分为2个相互密切联系的部分,即个体发育和系统发育,也就是个体的发育历史和由同一起源所产生的生物群的发展历史。
个体发育史是系统发展史的简单而迅速的重演。
”如青蛙的个体发育,由受精卵开始,经过囊胚、原肠胚、三胚层的胚,无腿蝌蚪、有腿蝌蚪,到成体青蛙。
这反映了它在系统发展过程中经历了像单细胞动物、单细胞的球状群体、腔肠动物、原始三胚层动物、鱼类动物,发展到有尾两栖到无尾两栖动物的基本过程。
说明了蛙个体发育重演了其祖先的进化过程,也就是个体发展简短重演了它的系统发展,即其种族发展史。
3 多细胞动物的起源-讲课

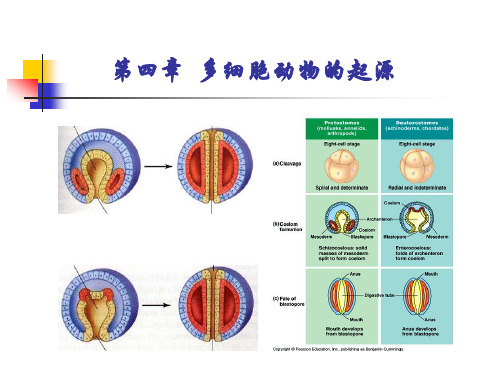
外胚层:分化为皮肤上皮(包括皮肤腺和其他皮肤 衍生物)、神经组织、感觉器官和消化管的两端。
三. 生物发生律
德国学者赫克尔(Haeckel, E. 1866)
生物发生律(biogenetic law ): 生物发展史分为个体发育和系统发育,即个体的发 育历史和由同一起源所产生生物群的发展历史。 个体发育史是系统发育史的简单而迅速的重演。 即生物在个体发育过程中,按顺序重演其祖先的主 要发育阶段。
不同地层的化石特点: 早期地层:古生物化石越简单,越低等, 晚期地层:古生物化石越复杂,越高等。 最明显、最主要的证据是化石。
最早的化石:澳州西海岸35亿年前的兰细菌化石
(二) 形态学证据(Morphological evidence)
现存的动物种类形成了一个由简单到复杂、由低等 到高等的序列。
辐射卵裂:棘皮、两栖类
第3次卵裂以后,上层的分裂球很整齐地排列在下 层之上,呈辐射排列。
不完全卵裂:多见于多黄卵,卵黄多,细胞分裂 受阻,卵裂只在不含卵黄的部位进行。
盘裂:分裂局限于胚盘处,如乌贼、鸡. 囊胚的形成(blastulation)
5. 中胚层及体腔的形成
中胚层的形成方式: 裂体腔法(端细胞法):原口动物、高等脊索;
体腔囊法(肠体腔法):棘皮动物、毛颚动物、半 索动物、原索动物。
体腔的类型:三胚层无体腔、假体腔和真体腔
6. 胚层分化
内胚层最简单,中胚层最复杂,外胚层最特异。
内胚层:分化为消化管的大部分上皮,肝、胰、呼 吸器官,排泄器官和生殖器官的一部分; 中胚层:分化为肌肉,结缔组织、生殖和排泄器官 的大部分;
4. 原肠胚的形成(Gastrulation)
第三章 多细胞动物的起源

蛙的大致发育过程 个体 受精卵 发育 系统 发育 单细胞 囊胚 单细胞群 原肠胚 腔肠动物 三胚层 三胚层动物 蝌蚪 鱼 有腿 有尾 两栖类 成蛙 无尾 两栖类
第五节 关于多细胞动物起源的学说
一、群体学说 大多数学者认为,多细胞动物起源于群体鞭毛虫类似的祖 先。对此也有两种假说: 1、原肠虫学说 赫克尔提出和团藻相似的群体单细胞动物一端内陷,形 成了有原肠和两胚层的原始多细胞动物。把此祖先称为原 肠虫。 2、吞噬虫学说 梅契尼可夫提出,具有单层细胞的单细胞群体内,一部 分细胞摄取食物后进入群体之内,形成了两胚层的实心的 原始多细胞动物。把此祖先称为吞噬虫。因现存的较低等 的动物,多由细胞移入而形成两胚层。内陷法到后来才有。 因此,吞噬虫学说可能更接近于事实。 二、合胞体学说 认为多细胞动物起源于多核纤毛虫的原始类群,后生动 物的祖先是具合胞体结构的多核细胞
第三章
多细胞动物的起源及 早期胚胎发育
第一节 从单细胞到多细胞
一、关于原生动物、中生动物和后生动物 二、关于多细胞动物的对称体制
第二节 多细胞动物起源于单细胞动物的证据
古生物学方面: 在距今愈古老的地层中,化石种类愈简单,在太古代 地层中,已发现有单细胞动物有孔虫的化石,而多细胞 动物化石在这里极少,说明单细胞动物的出现早于多细 胞动物。 形态学方面: 从群体鞭毛虫,如:盘藻、团藻等可以推测,群体是 单细胞动物过渡到多细胞动物的中间类型。 胚胎学方面: 多细胞动物的胚胎发育要经过受精卵、卵裂、囊胚、 原肠胚等阶段,均要经过单个细胞。根据“个体发育是 系统发育简短而迅速的重演”这一生物发生律,也说明 多细胞动物起源于单细胞动物。
第三节 胚胎发育的重要阶段
受精与受精卵 卵裂 囊胚的形成 原肠胚的形成 中胚层及体腔的形成 胚层的分化
第3、4章 多细胞动物的起源多孔动物门

第三章 多细胞动物的起源
从单细胞到多细胞
多细胞动物起源于单细胞动物的证据
关于多细胞动物起源的学说 胚胎发育的重要阶段 生物发生律
第四章 多孔动物门(Porifera)
多孔动物门的主要特征
分类及分类地位 经济意义
从单细胞到多细胞?
人的胚胎发育过程
动物的发育
个体发育 系统发育
• 一.个体发育:有机体从受精卵 发育成成体的过程。包括 胚前期, 胚胎期和胚后期。 (一)胚前发育:性细胞的产生和成熟的过程。
精母细胞 4个精子 3个极体
卵母细胞
1个卵细胞
少黄卵:卵黄少,均匀
中黄卵:卵黄多,分布于中央 多黄卵:卵黄多,分布于一侧
(二)胚胎发育
受精(fertilization)与受精卵 卵裂(cleavage) 囊胚的形成(blastulation) 原肠胚的形成(gastrulation) 中胚层及体腔的形成
活习性方面无区别,如低等昆虫。 变态发育(间接发育):幼体和成体在形态、生活 习性方面的区别显著,如蛙、昆虫等。
二、系统发育
个体发育和系统发育
个体发育(ontogeny): 是指生物个体从受精卵开始到个体性成熟 或死亡为止的整个过程。在个体发育过程中,个体的生理功能、
组织结构和器官形态都发生一系列变化。
生物发生律(biogenetic law, 重演律recapitulation law):
由德国科学家赫克尔(E.Haeckel)于1866年提出。是指个体发 育史是系统发展史的简单而迅速的重演。二者的对应关系如 下:
青 蛙
个 体 发 育
受 精 卵
囊 胚
第三章多细胞动物的起源

第3章多细胞动物的起源第1节从单细胞到多细胞一、知识点I、理论:一切高等动物虽然都是多细胞的,但其发展是不平衡的II、动物体复杂化的关键:对称体型和头部的形成III、两侧对称的意义:有利于动物活动;促使身体分为前后、左右、背腹IV、发展过程中3类动物:原生动物、中生动物、后生动物V、中生动物:一类小型的内寄主动物。
结构简单,分为菱形虫纲、直泳虫纲。
1、菱形虫纲:包括双胚虫、异胚虫。
无性生殖或有性生殖。
2、直泳虫纲:寄生在多种海生无脊椎动物体内。
成虫多雌雄异体,少数雌雄同体。
没有轴细胞。
VI、原始的多细胞动物:一般认为是中生动物,因为它和原声动物的纤毛虫类的亲缘关系比较近二、多细胞动物起源于单细胞动物的证据I、古生物学方面:古代动植物的遗体或残骸。
在最古老的地层中,化石种类是最简单的。
II、形态学方面:简单——>复杂;低等——>高等III、胚胎学方面:受精卵——>卵裂——>囊胚——>原肠胚第2节胚胎发育的重要阶段胚胎发育分为:受精与受精卵——卵裂——原肠胚的形成——中胚层及体腔的形成——胚层的分化I、受精与受精卵:精子与卵子结合为一个细胞称为受精卵II、卵裂:1、完全卵裂:多见于少黄卵。
a、等裂:海胆、文昌鱼;b、不等裂:海绵动物、蛙类。
2、不完全等裂:多见于多黄卵。
受精卵只在不含卵黄的部位进行分裂。
a、盘裂:乌贼、鸡卵;b、表面卵裂:昆虫卵III、囊胚的形成:囊胚:囊胚腔、囊胚层。
IV、原肠胚的形成:内陷、内移、内转、外包、分层。
最常见的是内陷和外包同时进行,分层和内移相伴而行V、中胚层及体腔的形成:端细胞法(裂体腔法);体腔囊法:棘皮动物、毛鄂动物、半索动物、脊索动物VI、胚层的分化:动物体的器官都是由内、中、外胚层发育而来1、内胚层:分化为消化管的大部分上皮、肝、胰、呼吸器官、排泄和生殖器官的小部分2、中胚层:分化为肌肉、结缔组织、生殖和排泄器官的大部分3、外胚层:分化皮肤上皮、神经组织、感觉器官、消化管的两端第3节生物发生律与多细胞起源学说一、生物发生律赫克尔在《普通形态学》中说:生物发展可分为2个密切联系的部分:个体发育和系统发展。
多细胞生物的起源
多细胞动物的起源姓名:王园学号:20117319专业:生物科学学院:农生院时间:2012.3.3随着科学的进步和人类的进一步探索,越来越多的例子和研究证明多细胞动物起源于单细胞动物。
在未来的日子里,揭开多细胞的起源将不会成为难题。
如:“多细胞生物起源或可追溯至寒武纪前”的发表证明人类的研究已经取得一定的成就,谜底解开之日将不会远。
一、多细胞动物起源于单细胞动物动物由单细胞演变为多细胞是动物发展史的一个重要阶段。
一切高等生物,包括动物、植物,都是多细胞的。
但多细胞动物的进化发展远较植物的快。
这是因为多细胞动物在进化过程中发展了两侧对称的体型,进而身体各部分明显分工,出现了头部,使得神经、感官等大大发展,而这些发展都是由于多细胞动物长期适应于活跃的、主动的生活方式而形成的。
原生动物——单细胞动物虽然也能完成起生命的各种活动,并有些单细胞动物结构上有一定程度的复杂化,但由各种细胞器来完成的各种不同的功能,这仅仅是一个细胞内的分化。
原生动物也有一些多细胞群体,它们只是以群体的方式存在,一般仍是以一个个细胞为独立的生活单位的,彼此之间并不发生密切联系。
多细胞动物我们称之为Metozoa,即后生动物,这是相对于原生动物(prot-ozoa)而言的。
后生动物包括除原生动物以外的绝大多数的多细胞动物。
也有学者认为在原生动物和后生动物之间存在着一个小类群--中生动物Mesozoa,这类动物寄生在海洋无脊椎动物的体内,个体细胞数目20-30个。
二、由单细胞动物发展到多细胞动物的证据现在公认多细胞动物起源与单细胞动物,证据主要如下三个方面:1、古生物学方面从不同地层中的化石种类来判断。
化石---古代动、植物的遗体或遗迹。
化石研究发现,越是古老的地层,化石种类越简单。
在太古代(地质史最古老的年代),距今32亿到距今18亿年的中生代的地层中有大量有孔虫的化石。
而晚近地层中的化石种类则较复杂,并且动物杂交的程度是一个渐进的变化过程。
多细胞动物的起源 笔记
第三章多细胞动物的起源第一节多细胞动物起源于单细胞动物的证据一、古动物学方面1、距今6亿年以前(古生代寒武纪)海相沉积里多为古老的原生动物化石(有孔虫,放射虫等),多细胞动物化石却很少见。
2、距今愈古老的化石中,种类愈少,愈简单;愈近的地层中,则愈多而复杂。
二、比较形态学方面三、胚胎学方面:多细胞动物是由受精卵开始,经过卵裂、囊胚、原肠胚等一系列过程,逐渐发育成成体。
第二节多细胞动物的系统发展绝大多数多细胞动物又叫后生动物,一般分为中生、侧生和真后生动物。
第三节多细胞动物的个体发育一、个体发育的概念:从受精卵或合子起到个体死亡的全过程。
二、三个阶段:1、性细胞的形成:增殖期:精原细胞和卵原细胞数量不断的增多时期;生长期:精原细胞——初级精母细胞;卵原细胞——初级卵母细胞成熟期:一个初级精母细胞——4个精子;一个初级卵母细胞——一个卵细胞和三个极体2、胚胎发育:从受精到幼体的产生,主要分以下阶段:(1)受精和受精卵:根据卵黄多少可将卵分为少黄卵、中黄卵和多黄卵。
卵黄相对多的一端称为植物极,另一端称为动物极。
精卵结合形成合子的过程称为受精。
受精方式有体内和体外两种。
(2)卵裂:定义:从受精卵经过多次的重复分裂,形成多个分裂球或胚泡的过程。
包括细胞的分裂、增殖和移位特点:分列的次数愈多,形成的分裂球愈小。
卵裂方式:完全卵裂:整个卵细胞都进行分裂,多见于少黄卵。
等裂:卵黄少,分布均匀,形成的分裂球大小相同,如文昌鱼;不等裂:卵黄在卵内分布不均与形成的分裂球大小不同:蛙类不完全卵裂:多见于多黄卵盘裂:分裂仅限于动物极胚盘上,乌贼和鸟类;表裂:分裂仅限于卵表面:昆虫卵(3)囊胚期:卵裂的结果,分裂球形成中空囊状的球状胚。
囊胚中间的腔称为囊胚腔。
囊胚壁的细胞层称为囊胚层。
(4)原肠期:胚胎分化出内胚层、外胚层、原肠腔、原口由囊胚植物极的细胞向内陷入。
最后形成2层细胞,在外面的细胞层称为外胚层,向内陷入的一层为内胚层。
多细胞动物的起源
胚胎发育的重要阶段
• 受精与受精卵 卵黄含量分 少黄卵、中黄卵、 按卵黄含量分:少黄卵、中黄卵、多黄卵 卵黄分布分 均黄卵、中黄卵、 按卵黄分布分:均黄卵、中黄卵、端黄卵 植物级与动物级:卵黄较多的一端称为植 植物级与动物级:卵黄较多的一端称为植 物级,另一端称为动物级 物级,另一端称为动物级。 动物级。 受精的生物学意义: 受精的生物学意义:子代能从亲代那里获 得双重的遗传性和变异性, 得双重的遗传性和变异性,使子代更具有生 命力。 命力。
胚层的分化
• 分化:同一类型细胞转变为复杂的、异质性和稳定性的 分化:同一类型细胞转变为复杂的、 细胞的过程。 细胞的过程。
各胚层的分化
• 外胚层:皮肤表皮、神经组织及感觉器官、消化管的两 外胚层:皮肤表皮、神经组织及感觉器官、 端。 • 中胚层:肌肉、结缔组织、生殖与排泄器官的大部分。 中胚层:肌肉、结缔组织、生殖与排泄器官的大部分。 • 内胚层:消化道大部分上皮,肝脏、胰脏,呼吸、排泄、 内胚层:消化道大部分上皮,肝脏、胰脏,呼吸、排泄、 生殖器官的小部分。 生殖器官的小部分。
关于多细胞动物起源的学说
• 群体学说
• Haeckel的原肠虫学说 : 囊胚虫 Haeckel的 ——→ 原肠虫 →
内移 内陷
• 梅契尼柯夫的吞噬虫学说:囊胚虫——→ 吞噬虫 梅契尼柯夫的吞噬虫学说:囊胚虫 吞噬虫学说 → • Grell-Butschli的扁囊胚虫学说: Grell-Butschli的扁囊胚虫学说: 扁囊胚虫 两侧原肠虫 ————————————————→ →
中生动物门:为一类小型的内寄生动物, 中生动物门:为一类小型的内寄生动物, 已知约50 已知约50种。分为菱形虫纲和直泳虫纲。 50种 分为菱形虫纲 直泳虫纲。 菱形虫纲和
多细胞动物的起源(1)
整理课件
21
第四节 关于多细胞动物起源的学说
(1)赫克尔原肠虫学说 最早的多细胞动物产生于类似
团藻的球形群体,这类群体一面 内陷形成具有双层细胞的多细胞 动物的祖先。这种祖先因为与原 肠胚相似,有两胚层和原口,赫 克尔称之为原肠虫(gastraea)
生物发生律是一条客观规律,它适用于整 个生物界。
不能把“重演”理解为简单机械的重复。个 体发育和系统发展是辩证统一的关系,系统发 展通过遗传决定个体发育,个体发育不仅简短 重演系统发展,而且也会出现新的变异,又能 补充和丰富系统发展。两种发育过程又都受环 境的影响。
整理课件
28
思考题:
1.中胚层形成的方式 2.原肠胚的特征
整理课件
3
第二节 多细胞起源于单细胞的证据
古生物学方面:最古老的地层中,化石种 类也是最简单的,而在晚近的地层中动物 化石种类也较复杂
形态学方面:从现有动物看,从单细胞动 物到多细胞动物形成了由简单到复杂,由 低等到高等的序列
胚胎学方面:根据生物发生律,及个体发 育简短地重演了系统发展的过程,可以说 明多细胞动物起源于单细胞动物
精卵细胞结合形成受精卵,是新个体的 开始,受精卵是单细胞
作用
使精卵细胞的单倍 染色体结合成合子 的双倍染色体
活化卵细胞
整理课件
7
二、卵裂(cleavage)
1、卵裂的概念、特点
• 精卵融合后,受精卵仍然是单个细胞,受精卵经 过多次分裂,形成很多分裂球的过程,称为卵裂
• 它是一种特殊的分裂方式,即在每次分裂之后, 未到新细胞长大就继续不断地分裂下去,因此分 裂成的细胞就越来越小,这些细胞叫做分裂球( blast onere)
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
多细胞动物的起源学说(一)群体学说大多数学者认为,多细胞动物起源于群体鞭毛虫类似的祖先。
对此也有两种假说:1、原肠虫学说赫克尔提出和团藻相似的群体单细胞动物一端内陷,形成了有原肠和两胚层的原始多细胞动物。
把此祖先称为原肠虫。
2、吞噬虫学说梅契尼可夫提出,具有单层细胞的单细胞群体内,一部分细胞摄取食物后进入群体之内,形成了两胚层的实心的原始多细胞动物。
把此祖先称为吞噬虫。
因现存的较低等的动物,多由细胞移入而形成两胚层。
内陷法到后来才有。
因此,吞噬虫学说可能更接近于事实。
(二)合胞体学说认为多细胞动物起源于多核纤毛虫的原始类群,后生动物的祖先是具合胞体结构的多核细胞。
生物发生率(biogenetic law)也叫重演律(recapitulation law),是德国人赫克尔(E.H.Haeckel)用生物进化论的观点总结了当时胚胎学方面的工作提出来的。
当时在胚胎发育方面已揭示了一些规律,如在动物胚胎发育过程中,各纲脊椎动物的胚胎都是由受精卵开始发育的,在发育初期极为相似,以后才逐渐变得越来越不相同。
达尔文曾作过一些论证,认为胚胎发育的相似性,说明它们彼此有亲缘关系,起源于共同的祖先,个体发育的渐进性是系统发展中渐进性的表现。
达尔文还指出了胚胎结构重演其过去祖先的结构,“它重演了它们祖先发育中的一个形象”。
海克尔明确地论述了生物重演律。
1866年他在《有机体普通形态学》书中说:“生物发展史可分为2个相互密切联系的部分,即个体发育和系统发育,也就是个体的发育历史和由同一起源所产生的生物群的发展历史。
个体发育史是系统发展史的简单而迅速的重演。
”如青蛙的个体发育,由受精卵开始,经过囊胚、原肠胚、三胚层的胚,无腿蝌蚪、有腿蝌蚪,到成体青蛙。
这反映了它在系统发展过程中经历了像单细胞动物、单细胞的球状群体、腔肠动物、原始三胚层动物、鱼类动物,发展到有尾两栖到无尾两栖动物的基本过程。
说明了蛙个体发育重演了其祖先的进化过程,也就是个体发展简短重演了它的系统发展,即其种族发展史。
生物重演律对了解各动物类群的亲缘关系及其发展线索极为重要。
因而对许多动物的亲缘关系和分类位置不能确定时,常由胚胎发育得到解决。
生物重演律是一条客观规律,它不仅适用于动物界,而且适用于整个生物界,包括人类在内。
简而言之,生物发生率揭示了个体发育是系统发育快速的重演群落 community 亦称生物群落(biological community)。
生物群落是指具有直接或间接关系的多种生物种群的有规律的组合,具有复杂的种间关系。
我们把在一定生活环境中的所有生物种群的总和叫做生物群落,简称群落。
组成群落的各种生物种群不是任意地拼凑在一起的,而有规律组合在一起才能形成一个稳定的群落。
如在农田生态系统中的各种生物种群是根据人们的需要组合在一起的,而不是由于他们的复杂的营养关系组合在一起,所以农田生态系统极不稳定,离开了人的因素就很容易被草原生态系统所替代。
结构任何群落都有一定的空间结构。
构成群落的每个生物种群都需要一个较为特定的生态条件;在不同的结构层次上,有不同的生态条件,如光照强度、温度、湿度、食物和种类等。
所以群落中的每个种群都选择生活在群落中的具有适宜生态条件的结构层次上,就构成了群落的空间结构。
群落的结构有水平结构和垂直结构之分。
群落的结构越复杂,对生态系统中的资源的利用就越充分,如森林生态系统对光能的利用率就比农田生态系统和草原生态系统高得多。
群落的结构越复杂,群落内部的生态位就越多,群落内部各种生物之间的竞争就相对不那么激烈,群落的结构也就相对稳定一些。
群落有其结构。
大多数群落中,由一两种占优势的植物生长型决定整个群落的外貌,群落也常以此得名,如阔叶落叶林、针叶常绿林、草原等。
植物还可以按更新芽的位置而分为不同生活型,如地上芽、地下芽植物等。
一个群落的生活型组成可以反映环境特征。
群落还常表现垂直分层现象,如地面上高树、矮树、灌木、草本的分层与光照有密切关系。
地下和水中生物亦如是。
除光照外,氧气、压力等亦有关。
以植物为栖息地和食物的动物亦有相应的分层。
在水平方向,不同生物可因要求类似环境条件或互相依赖而聚集在一起。
群落中各物种常随时间而变化,如植物的开花闭花和动物的穴外行动具有昼夜节律,而整个温寒带群落呈现明显季节节律。
群落中生物总处在不断的交互作用中。
按生物吸取营养的方式,有营光合作用的植物、靠摄食为生的动物和经体表吸收的微生物。
它们之间形成复杂的食物关系。
两物种可以是互相竞争,也可是共生,视相互间利害关系而有寄生、偏利共生和互利之分。
一个群落的进化时间越长、环境越有利且稳定,则所含物种越多。
如两物种利用相同资源(生态位重叠)则必然竞争而导致一方被排除。
但如一方改变资源需求(生态位分化)则可能共存。
生物群落的发展趋势是生态位趋向分化和物种趋向增多。
植物通过光合作用制造的有机物质总量称为总初级生产力,这是整个群落一切生命活动的能量基础。
除去植物呼吸消耗之後的剩馀称为净初级生产力,这是群落中全部异营生物(亦称异养生物)赖以生存的能源。
群落中现存的有机物质量称为生物量,各种类型的群落的生物量和生物量积累比率很不相同。
群落中生物组成包括植物、食植动物到食肉动物各营养级的食物连锁关系。
由于能量的种种消耗,生产力逐级递减。
初级生产力只占阳光能中的0.1~1%,而动物所代表的各次级生产力只占前一级生产力的10%。
土壤上下的细菌、真菌在群落中亦占重要地位。
森林中被动物摄食者,不到枝干量的1%和树叶量的10%,绝大部分朽木落叶被微生物分解。
有机物质被分解为简单成分後,可再为根系所利用从而完成营养物循环。
森林中这种循环可以很紧密,丢失很少。
但海洋中浮游生物沉积海底,却使一部分营养物(如磷)难以再重复利用。
一片山坡上的丛林可因山崩全部毁坏,暴露出岩石面。
但又可经地衣、苔藓、草类、灌木和乔木等阶段逐步再发育出一片森林,包括重新孕育出土壤。
当一个群落的总初级生产力大于总群落呼吸量,而净初级生产力大于动物摄食、微生物分解以及人类采伐量时,有机物质便要积累。
于是,群落便要增长直达到一个成熟阶段而积累停止、生产与呼吸消耗平衡为止。
这整个过程称为演替(succession),而其最後的成熟阶段称为顶极(climax)。
顶极群落生产力并不最大,但生物量达到极值而净生态系生产量很低或甚至达到零;物种多样性可能最後又有降低,但群落结构最复杂而稳定性趋于最大。
不同于个体发育,群落没有个体那样的基因调节和神经体液的整合作用,演替道路完全决定于物种间的交互作用以及物流、能流的平衡。
因此顶极群落的特征一方面取决于环境条件的限制,一方面依赖于所含物种。
垂直结构形成原因:群落中,各个生物种群分别占据了不同的空间。
概念:垂直结构是指在群落生境的垂直方向上,群落具有的明显分层现象。
以森林的群落结构为例。
在植物的分层上,由上至下依次是乔木层、灌木层和草本植物层。
动物的分层亦呈这种垂直结构:鸟类分为林冠层,中层和林下层。
林冠层包括鹰,伯劳,杜鹃,黄鹂等。
中层包括山雀,莺,啄木鸟等。
林下层包括画眉,八色鹊等。
水体分层也是如此。
水体分为上层,中层和底层。
上层主要是藻类。
中层主要为浮游动物。
底层主要为软体动物,环节动物和蟹类。
水平结构水平结构是指在群落生境的水平方向上,群落具有的明显分层现象。
由于在水平方向上存在的地形的起伏、光照和湿度等诸多环境因素的影响,导致各个地段生物种群的分布和密度的不相同。
同样以森林为例。
在乔木的基部和被其他树冠遮盖的位置,光线往往较暗,这适于苔藓植物等喜阴植物的生存;在树冠下的间隙等光照较为充足的地段,则有较多的灌木与草丛。
分类生态学研究中常将群落分类并加以排序,但因物种单独适应环境而群落间是逐渐过渡,故分类缺乏明确界线。
选择不同分类标准得出不同结果。
一般生物群落分类藉用植物群落分类系统。
详细研究特定地区内的植物群落,常以群丛为基本单位,根据特征种定出群丛,再顺次组成群属、群目、群纲等。
在大陆范围上,则主要按优势顶极画分成不同生物群系,它们反映不同的气候地质条件。
常见群系类型如海洋、淡水、沼泽、森林、荒漠、冻原等等。
热带雨林分布在高温多雨的热带地区。
物种丰富,层次多,最复杂。
热带雨林主要分布于赤道南北纬 5 ~ 10度以内的热带气候地区。
这里全年高温多雨,无明显的季节区别,年平均温度 25 ~30 ℃,最冷月的平均温度也在18 ℃ 以上,极端最高温度多数在36 ℃ 以下。
年降水量通常超过 2 000mm ,有的竟达 6 000mm ,全年雨量分配均匀,常年湿润,空气相对湿度 90 %以上。
热带雨林为热带雨林气候及热带海洋性气候的典型植被。
大多数热带雨林(Tropical zone rain forest)都位于北纬23.5度和南纬23.5度之间。
在热带雨林中,通常有三到五层的植被,上面还有高达150英尺到180英尺的树木像帐篷一样支盖着。
下面几层植被的密度取决于阳光穿透上层树木的程度。
照进来的阳光越多,密度就越大。
热带雨林主要分布在南美、亚洲和非洲的丛林地区,如亚马逊平原和云南的西双版纳。
每月平均温度在华氏64.5度以上(摄氏温度约为18度),平均降水量每年80英寸(1英寸=2.54厘米)以上,超过每年的蒸发量。
常绿阔叶林分布在温暖多湿的亚热带地区。
常绿阔叶林是亚热带海洋性气候条件下的森林,大致分布在南、北纬度22°~34°(40°)之间。
主要见于亚洲的中国长江流域南部、朝鲜和日本列岛的南部,非洲的东南沿海和西北部,大西洋的加那利群岛,北美洲的东端和墨西哥,南美洲的智利、阿根廷、玻利维亚和巴西的部分地区,大洋洲东部以及新西兰等地。
其中以中国长江流域南部的常绿阔叶林最为典型,面积也最大。
由常绿阔叶树种组成的地带性森林类型。
针叶林(taiga forest regions)分布:寒温带及中、低纬度亚高山地区植物:冷杉,云杉,红松热带草原(savanna or savannah)分布:干旱地区。
特点:年降水量少,群落结构简单,受降雨影响大;不同季节或年份种群密度和群落结构常发生剧烈变化,景观差异大。
荒漠(desert)分布:南北纬15°~50°之间的地带。
特点:终年少雨或无雨,年降水量一般少于250mm,降水为阵性,愈向荒漠中心愈少。
气温、地温的日较差和年较差大,多晴天,日照时间长。
风沙活动频繁,地表干燥,裸露,沙砾易被吹扬,常形成沙暴,冬季更多。
荒漠中在水源较充足地区会出现绿洲,具有独特的生态环境。
冻原(tundra)分布:欧亚大陆和北美北部边缘地区,包括寒温带和温带的山地与高原。
特点:冬季漫长而严寒,夏季温凉短暂,最暖月平均气温不超过14℃。
年降水200~300mm。
沼泽分布于低洼地和排水不良地段,可分为草本沼泽和森林沼泽静息电位静息电位(Resting Potential,RP)是指细胞未受刺激时,存在于细胞膜内外两侧的外正内负的电位差。